
次はディスコセア綱(class Discosea)に属する Acanthamoeba castellanii ペントースリン酸経路である。
Acanthamoeba castellanii はアメーバ型の原生生物の一種で、淡水域やその他の場所にも広く分布する土壌微生物である。アカントアメーバ属の大半はバクテリアを捕食して生活する従属栄養生物であるが、一部の種は感染性を持ち、ヒトや他の動物に対して角膜炎や脳炎(アメーバ性肉芽腫性脳炎)を引き起こす。このAcanthamoeba castellanii もアカントアメーバ角膜炎を引き起こすらしい。とはいえ、Acanthamoeba castellanii の持つペントースリン酸経路はいままで何度も見てきたものであり、NADPH2のみならず完全な形の糖相互変換系を持つオーソドックスなものでありこれ以上の議論は不要であろう。
次は原生生物の大きなグループであるアピコンプレックス門(Apicomplexa;またはアピコンプレクサ類)に属する原生動物についてである。アピコンプレックス門の下位分類については分子系統解析による再検討が行われている段階である。Aconoidasida (無コノイド綱)は分子系統解析の導入以前に微細構造に基づき設定されたものであるが、主要な生物群においては分子系統解析でも比較的安定的なクレードを形成する。おせっかいかもしれないが、クレードとは共通の祖先から種分化により派生した子孫のすべてを含む種の1群を意味する分岐群をいう。アピコンプレクサ類は、生活環のどこかでアピカルコンプレックス(apical complex、頂端複合構造)という構造を持つ原生生物の大きなグループである。寄生性であり、配偶子の時期以外は鞭毛や仮足を持たない。
| Apicomplexa (apicomplexans) |
| Aconoidasida |
| Haemosporida |
| pfa Plasmodium falciparum 3D7 |
| pfd Plasmodium falciparum Dd2 |
| pfh Plasmodium falciparum HB3 |
| prei Plasmodium reichenowi |
| pgab Plasmodium gaboni |
| pyo Plasmodium yoelii |
| pcb Plasmodium chabaudi |
| pbe Plasmodium berghei |
| pvv Plasmodium vinckei vinckei |
| pkn Plasmodium knowlesi |
| pvx Plasmodium vivax |
| pcy Plasmodium cynomolgi |
| Piroplasmida |
| tan Theileria annulata |
| tpv Theileria parva |
| tot Theileria orientalis |
| beq Theileria equi |
| bbo Babesia bovis |
| bmic Babesia microti |
| bbig Babesia bigemina |
| Conoidasida |
| Eucoccidiorida |
| tgo Toxoplasma gondii |
という事で、まず Haemospororida(住血胞子虫)から始めよう。住血胞子虫は赤血球や白血球に寄生する原生生物の一群で、代表的なものはマラリア原虫である。陸上脊椎動物を中間宿主、吸血昆虫を終宿主とする場合が多い。 Plasmodium falciparum 3D7から Plasmodium cynomolgiまで、12種のマラリア病原体類は、ほぼ同じペントースリン酸経路を持つ。
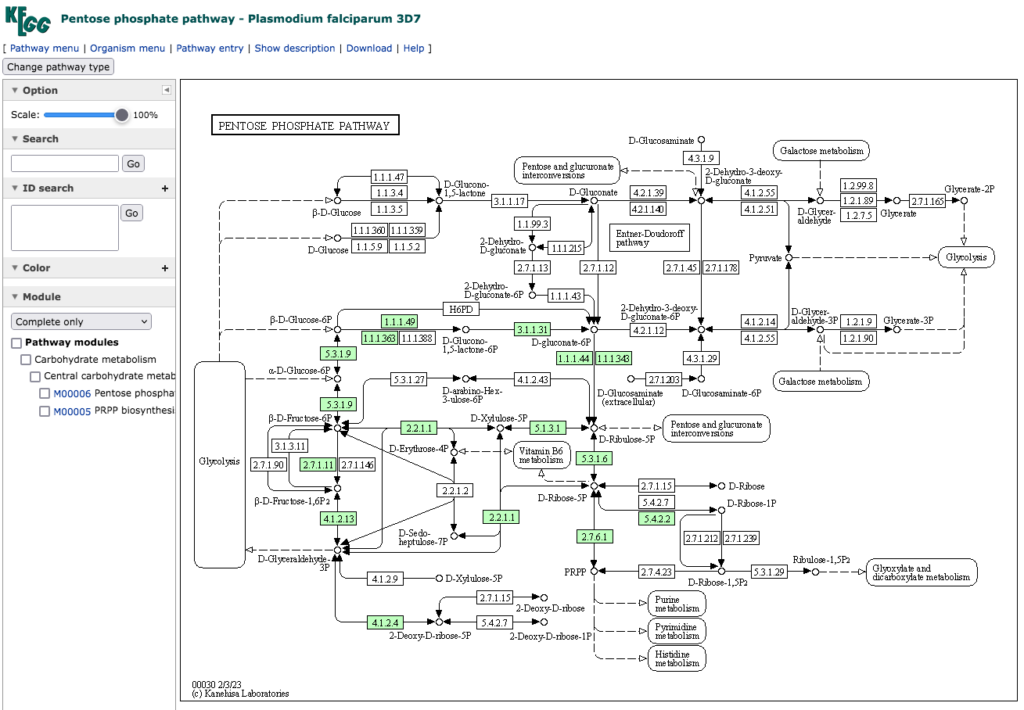
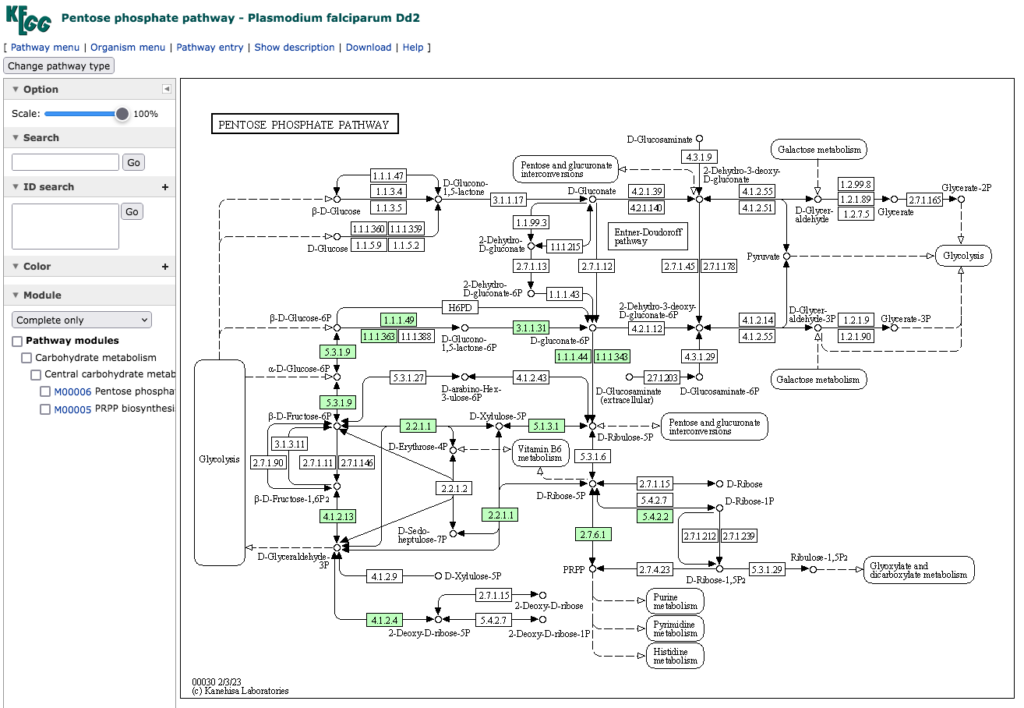
下段にPlasmodium falciparum Dd2のペントースリン酸経路を示している。それ以外の11種のマラリア病原体は、上段に示した Plasmodium falciparum 3D7 の持つペントースリン酸経路と同じものを持っている。この系を見れば、NADPH2 生合成もできているしリボース−5−リン酸の生合成も機能している。共生微生物であるにもかかわらず大事な遺伝子は残しているという解釈で良いだろう。ただ、全ての種においてペントースリン酸経路の華とでも言うべき酵素(個人的感想にすぎない)であるTransaldolase (EC 2.2.1.2:Sedoheptulose-7-phosphate:D-glyceraldehyde-3-phosphate glyceronetransferase )を欠いているのは興味深い。この酵素が無くてもペントースリン酸経路は機能するのである。この酵素の反応を有機電子論的にトレースできず、長期間悩んだことを思い出している。
下段に示したPlasmodium falciparum Dd2 の持つペントースリン酸経路 においては、Ribose phosphate isomerase(EC 5,3, 1,6: D-ribose-5-phosphate aldose-ketose-isomerase)に触媒される反応が欠損している。EC 2.2.1.2と同時にEC 5,3, 1,6を欠いた場合、リボース–5–リン酸の生合成が上手くできないのではないかと思うのだが、この反応は酵素が無くても少しづつ進むのではないかと思われる。
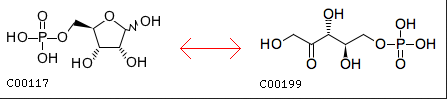
水中で、リブロース–5−リン酸(上図右)の1位と2位の間でケト–エノール異性化が起こり1位の炭素がアルデヒドとなった後、この炭素原子を2位の水酸基が攻撃して閉館すれば左のリボース−5−リン酸が生成する反応は、糖化学においてはよく起こる反応である。両化合物の水中における平衡定数を精査すべきだとは思うが、さほどの問題が起きない可能性はあるだろう。
次はアピコンプレックス門無コノイド綱ピロプラズマ目の Theileria 科とBabesia 科の生物である。過去からの分類学の流れで、小型のタイレリア科と大型のバベシア科に分けてはいるが、これは生物の系統を反映しておらず Babesia 属や Theileria 属は多系統的であることが明らかになっている。両者ともにマダニを終宿主とする寄生虫で、吸血された哺乳動物や鳥類の血球に寄生する原生生物である。 熱帯において家畜に対する致死的な病原体として警戒されているが、日本でも放牧牛の間で蔓延している小型ピロプラズマ症病原体も含まれている。
ペントースリン酸経路の構成から見ると、Theileria 科の4種とBabesia bovis が下に示した同じ経路を持っている。
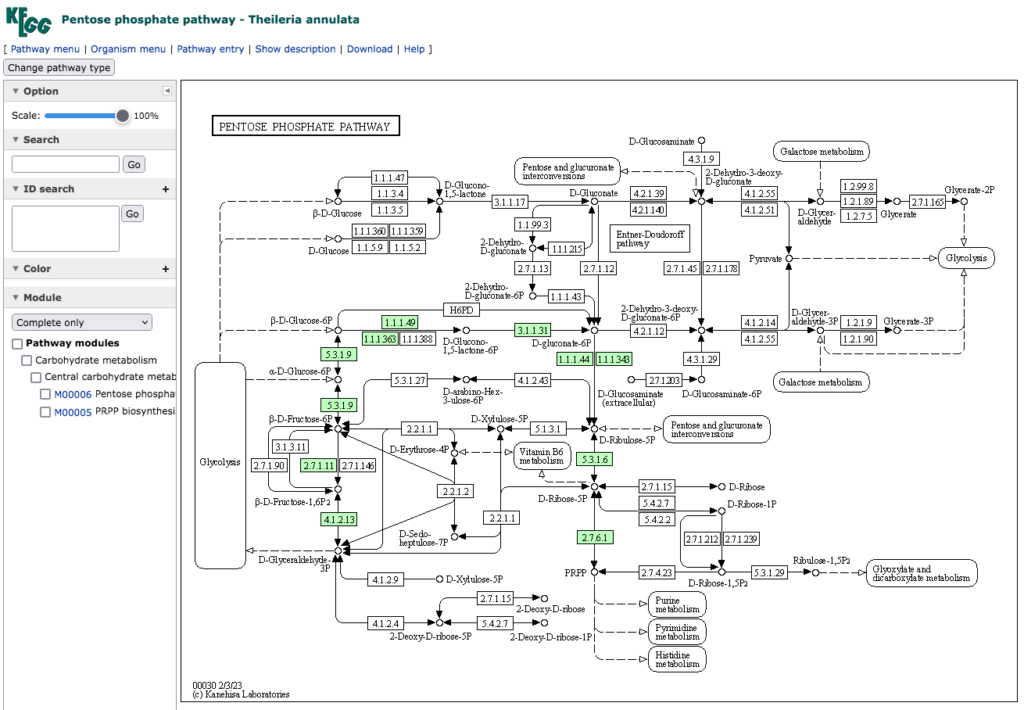
先に述べた Transaldolase (EC 2.2.1.2:Sedoheptulose-7-phosphate: D-glyceraldehyde-3-phosphate glyceronetransferase )を欠いているだけでなく、3単糖、4単糖、5単糖、6単糖、7単糖の相互変換を担っている経路が抜け落ちている。にもかかわらず、NADPH2 の生合成もできているしリボース−5−リン酸の生合成も機能している。あの理解が難しかったペントースリン酸経路が、その存在意義を維持したままこんなに簡素化できるのかと驚いている。とはいえ、ペントースリン酸経路を、解糖系の一部とあわせて回路を形成していると見なしペントースリン酸回路と称する場合があるが、これら生物のペントースリン酸経路は最早回路を形成していないため回路と云う用語は適切ではないようだ。
次は Babesia microti の持つペントースリン酸経路である。
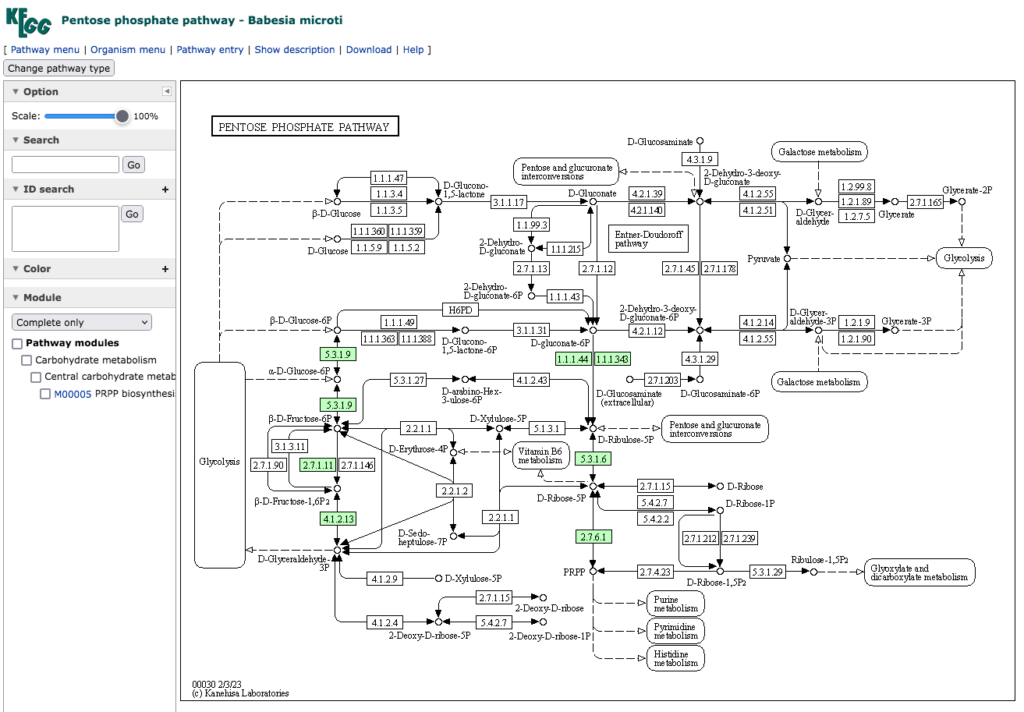
この原虫の主要な病原体保有生物は齧歯類であり、通常はマダニ科のシカダニが媒介する。ダニ幼虫は感染を受けた齧歯類を吸血して感染し、変態して若虫となり、原虫を他の動物またはヒトに伝播する。詳しくはhttps://www.msdmanuals.com/ja-jp/%E3%83%97%E3%83%AD%E3%83%95%E3%82%A7%E3%83%83%E3%82%B7%E3%83%A7%E3%83%8A%E3%83%AB/13-%E6%84%9F%E6%9F%93%E6%80%A7%E7%96%BE%E6%82%A3/%E8%85%B8%E7%AE%A1%E5%A4%96%E5%AF%84%E7%94%9F%E5%8E%9F%E8%99%AB/%E3%83%90%E3%83%99%E3%82%B7%E3%82%A2%E7%97%87 を参照のこと。この原虫の持つペントースリン酸経路 においては、NADPH2 の生産のみならずいわゆる解糖系との接続も切れてしまっており、経路としての体をなしていない。これを見てどう解釈すればいいのか、途方に暮れるしかない。寄生生物であると云うロジックに頼るしかなさそうだ。
次はBabesia bigemina についてだが、牛、水牛および鹿に感染し、中南米、東南アジア、アフリカ、豪州に分布し、オウシマダニをはじめとするコイタマダニ属のマダニによって媒介される。これまた詳しくはhttps://www.naro.affrc.go.jp/org/niah/disease_fact/k13.htmlを参照して下さい。
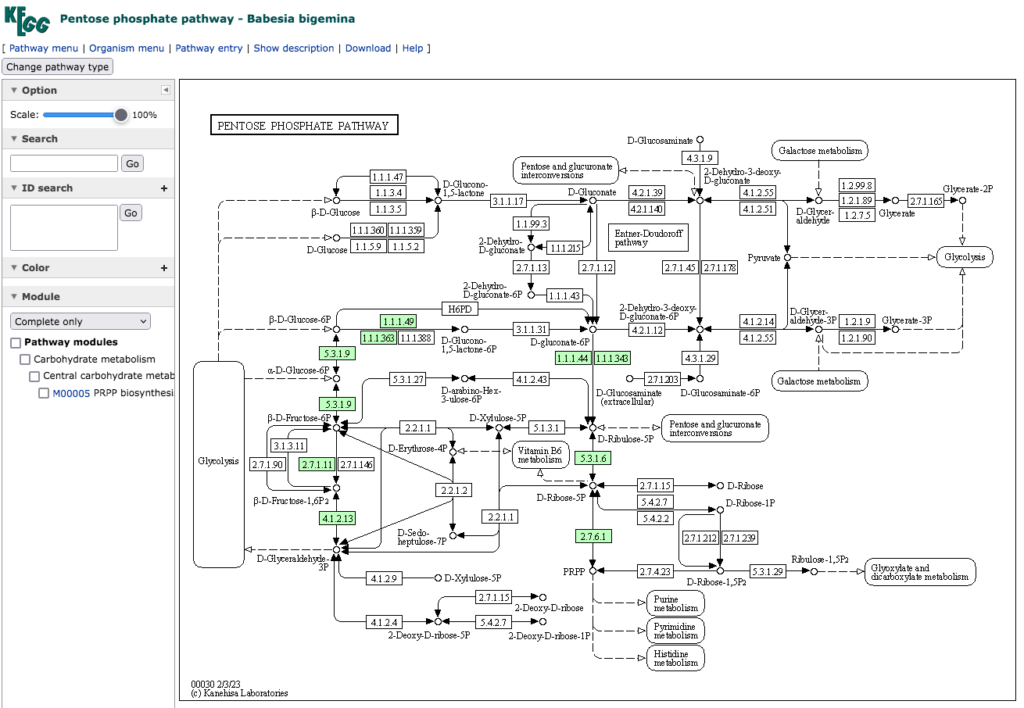
この原虫においては 6-phosphogluconolactonase(EC 3.1.1.31)を欠いていて、一見二つの系が繋がっていないように見えるのだが、この酵素は次の反応を触媒する。
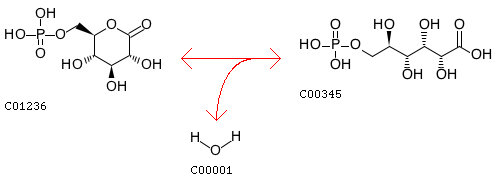
都合の良い解釈ばかり持ち出して気が重いのだが、これは単なる分子内環化反応であり水中であっても進まないわけではない。他の反応で機能する基質特異性の低い脱水酵素が、この段階を担っているかもしてないし、ホストのもつ酵素を借用している可能性も捨てきれない。などというでたらめに近い言い訳で NADPH2 生合成は可能かもしれないということにしておこう。言うまでもなく、そのスピードは高くはないに違いない。Babesia bigemina についての詳しい情報は下記を参照のこと。
https://www.rakuno.ac.jp/wp-content/themes/rgu/file/reserch-report_1212.pdf
今回は何となく自信を持てない。共生と云う現象に頼りすぎているのかもしれない。でもここを通らないと次にいけない気がしています。とにかく中間段階にあるという事です。