
歳を重ねると物事の処理速度が大きく低下するにもかかわらず、処理すべき事柄が減るわけではない。年寄りが忙しい忙しいと言うわりにはノロノロしているように見える理由であろう。これは歳をとってみないとわからない。これに痴呆が被ってくると事はさらに悲惨な事になってくるに違いない。何でこんなことを書くかと言えば、私自身が明らかな老化の足音を聴く年代になったからである。
前回のアップからどれくらい経ったか記憶にないが、こうして再び書き始めようとすると何をどう議論しようとしていたのか、記憶は薄れてしまいはなはだ心許ない。読者の年齢は知る術もないが、書いている本人の記憶がそうなのだから読者が以前の記事を記憶していると期待する方が図々しいだろう。従って、記憶がしっかりしているであろう若者には不要かもしれないが、何故にペントースリン酸経路に対して異論を唱える事になったか、手短に述べる事から始める事にする。
ことの初めは解糖系におけるブドウ糖の位置づけについての疑問であった。解糖系に対し一つの疑問を抱いた事が発端である。解糖系は大腸菌からヒトまでどころではなく、原核生物(古細菌を含む)から真核生物にわたるほとんどの生物に広く分布する普遍的な代謝系と理解されている。その点について大きな疑問はないのだが、出発物質がブドウ糖であることに納得が行かなかった。解糖系の起源を考える場合、現在の常識に従えばブドウ糖がアプリオリに存在する前提で書かれていると考えざるを得ないが、その場合アプリオリに存在していたブドウ糖の起源は何処あったのか。ホルムアルデヒドが粘土鉱物の表面でアルドール縮合を続ければ、D-ブドウ糖が生成する可能性がない事もないが、だからといってD-ブドウ糖が特異的に集積する確率は絶望的に低いと考える。さらに、解糖系らしき系は持つものの、その系の中にブドウ糖を含まない生物も少なからず散見される。それらをどのように説明すればいいのか?
私なりの答えは「歴史生物学・解糖系についての考察1〜5」に書いているので、その部分を参照して欲しい。系の起源の問題として捉えれば、解糖系に寄り添っている糖新生系の方がより古くより根源的であると言う結論にならざるを得ない。そう考えると、言い過ぎである事は自覚しているが、グルコースは解糖系からはみ出した盲腸であるとする考えさえ成立する。
だが、世の中では解糖系こそが生物に存在する多くの代謝系のなかで非常に重要な経路であり、中心経路と呼ばれる立ち位置にあるとする考え方が主流である。その解糖系の意義付けが変わると、解糖系に連接する経路の存在意義に変化が生じるかもしれないと云う問題意識を持ってTCA回路を眺めてみた。
そういう意識のもとにTCA回路を眺めると、解糖系の場合と同じように逆向きに反応が進行する嫌気的カルボン酸サイクルの存在とその存在意義がいま一つ曖昧なまま残置されている事に気付いた。さらに、現在行われているTCAサイクルの意義付けにおいては、α–ケトグルタル酸からのアミノ酸・タンパク質代謝に言及しているとは言え、TCA回路に続く電子伝達系と組み合わせてエネルギー獲得に重きを置く説明が優位な状況にある。
歴史生物学的視座から考える場合、ある系路の原初的な形はどのようなものであったかを問題とする。勿論、過去の生物そのものがもっていた回路をそのまま調べるわけには行かないが、過去の回路を現在も維持していそうな生物種における代謝系から予想できない事はない。 さて、アセチルCoAがオギザロ酢酸と縮合してクエン酸となるのだが、回路の出発点をオギザロ酢酸におきクエン酸を通って α–ケトグルタル酸に至る右回りの系路(東廻り系路)とリンゴ酸を通ってα–ケトグルタル酸に至る左回りの系路(西廻り系路)に分けてみた。そうすると結構面白い結果が得られる。 例えば、Methanothermococcus やMethanocaldococcus属細菌は西廻りの系路を持つが東廻りの系路は持たない。
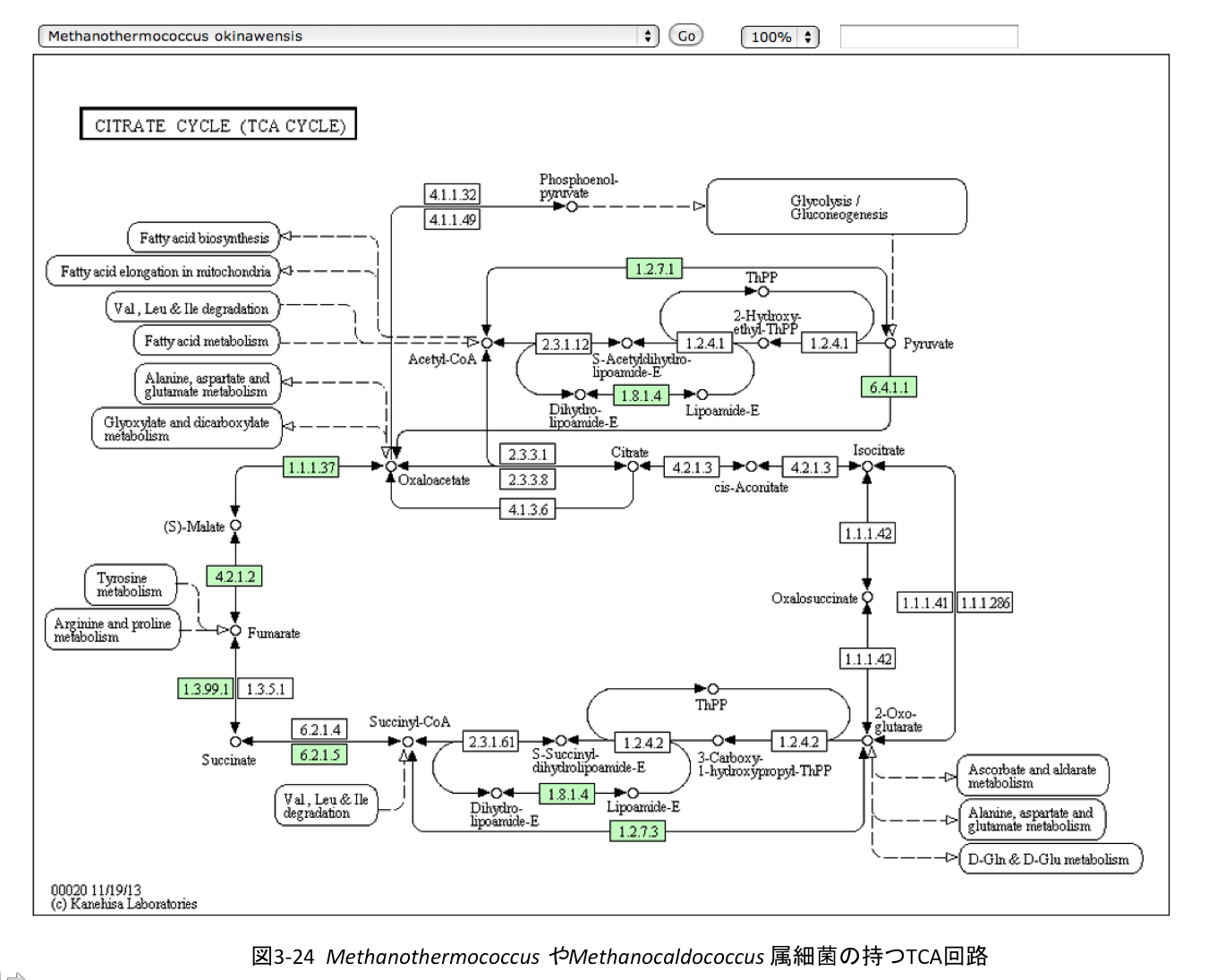
この西廻り系路を持つ生物は古細菌だけではない。真性細菌であるChlamydia、Chlamydophil、Bifidobacterium 、Prevotella、Desulfotomaculumも、同じようにオギザロ酢酸からα-ケトグルタル酸までの西廻りの代謝を行い東廻りの系路は持たない。
一方、TCA回路の西廻りの系を持たず東廻りの系のみを持つ生物も種々存在する。Eubacterium、Roseburia、Coprococcus、 Ruminococcus 属細菌などである。
 この現実を基礎に考えれば、TCA回路を回路と命名することの不合理さとともに、系の意義をエネルギー(ATP)生産系とする説明は現実を正しく反映しているとは思えない。幾分強引な推論だが、いわゆる解糖系と還元的カルボン酸サイクルに加えて、図3-24と図3-25の系について考えれば良さそうだ。これらの4種の回路と・非回路を持つそれぞれの生物群において、それぞれの系は同じ目的で駆動されていると考えて良いだろう。幾つかの異なるしかしよく似た代謝系が同じ目的を持って駆動している場合、まず各集団に共通するものを探すのが鉄則である。この場合、4つの系で共通している化合物は α-ケトグルタル酸である。
この現実を基礎に考えれば、TCA回路を回路と命名することの不合理さとともに、系の意義をエネルギー(ATP)生産系とする説明は現実を正しく反映しているとは思えない。幾分強引な推論だが、いわゆる解糖系と還元的カルボン酸サイクルに加えて、図3-24と図3-25の系について考えれば良さそうだ。これらの4種の回路と・非回路を持つそれぞれの生物群において、それぞれの系は同じ目的で駆動されていると考えて良いだろう。幾つかの異なるしかしよく似た代謝系が同じ目的を持って駆動している場合、まず各集団に共通するものを探すのが鉄則である。この場合、4つの系で共通している化合物は α-ケトグルタル酸である。
ではこの α-ケトグルタル酸、生物の中でどのような意義を持つのか。多くの方がご存知の通り、α-ケトグルタル酸からはアミノ酸代謝が出発する。α-ケトグルタル酸の α-位のカルボニル基にアミノ基転移が起こりL-グルタミン酸、L-グルタミン酸の γ 位のカルボキシル基がアミド化されてL-グルタミンがつくられた後、これらが種々の α-ケト酸にアミノ基を転移して、多様なアミノ酸類の生合成が起こる。アミノ酸生合成は、生物がタンパク質をつくる前段階の欠くべからざる反応であり、「その出発物質であるL-グルタミンとL-グルタミンの原料である α-ケトグルタル酸を供給するのがTCA(非)回路の意義である」とするのが適切な判断だと思う。
こんなブログを読んでいる方であれば、α-ケトグルタル酸から誘導されたL-グルタミン酸とL-グルタミンの存在意義は、先に述べたアミノ酸生合成の出発点であるだけではない。両化合物は、プリン代謝につながりDNAとRNAの原料であるアデニン、グアニンの生合成につながるのみならず、ピルビン酸の脱炭酸段階で必要なチアミンリン酸生合成へと伸びている。それだけではない。両化合物はピリミジン環生合成の原料でもありDNAとRNAの原料であるチミン、ウラシル、シトシンへと伸びるの生合成を可能にしている。
つまり、α-ケトグルタル酸はアミノ酸生合成とタンパク質生合成の中核に位置する化合物であると同時に、プリン塩基とピリミジン塩基生合成担う共通の原料でもある。これらの事実を重ね合わせてこの系の意義を求めるとすれば、第一義的には α-ケトグルタル酸の供給という役割を見なければならない。酸化的リン酸化系を持つミトコンドリアの祖先となる好気性細菌が原真核生物と共生した際に、東廻りのTCA回路が酸化的リン酸化と共役して多量のATPの生産を始めたのは、大気中の酸素濃度が上昇した後の話であろう。
ことの詳細については、歴史生物学 TCA回路への異論 1〜9に書いている。
そこで目下の問題は、ペントースリン酸経路についての解釈である。実は何人かの大学教授の方々にペントースリン酸経路の意義について尋ねたたことがあるのだ、上手く説明する人に会った事がない。解糖系の副路でNADPH2の生合成系路だったよねという教科書的な答えが帰ってきた。あれは複雑で分かり難いとか授業をやり難い系路だと云う本音のような愚痴のような答えもあった。同感である。私も不惑の頃まで、この部分の講義をする時は、前夜から予習をしていても落ち着かなかった。
ペントースリン酸経路に対して、糖代謝の根幹を担う解糖系と糖新生系に纏わりつく藤の木みたいに感じていた。天皇家に巻き付いた藤原一族といえば、感覚的に分かり易いかもしれない。先に述べたように、この解糖系・糖新生系の存在解釈に変更が生じたとすれば、この寄生植物のようなペントースリン酸経路の存在意義も影響を受けるに違いない。そう思って書いてきたのが、ペントースリン酸経路への異論1〜6である。そこに多分間違いないと思える答えは書いている。本人の中では結論が出ているのだが、文書の形を整えるためには、いま少しの考察を続ける必要が残っている。それは理解しているのだが、そこに手間ひまかけるのは精神的に辛いものがある。
論文を執筆する場合、結果は見えていると思えるにもかかわらず、いくつかの穴埋めのために補充実験が必要になる場合がある。レフェリーは必ずその穴を指摘して書き直しを命じてくる。この補充実験が無駄に思えて何度取り下げた事か。結果は見えているので次の実験に取り掛かりたいのに、何で時間の無駄とも思える穴埋のための補充実験に戻らなければならないのかと、我が侭である事は分かりながらも逃避したわけだ。まあ実験計画の不備というのが現実であり、私の責任であるから、レフェリーに罪はない。穴埋め実験を学生に強制できるのが教授の特権だと云う人もいるくらいだから、誰にでも発生する事案であろう。
先にも述べたように全種類の生物のペントースリン酸経路を順に検討するのはコストパフォーマンスが悪いので、次回からは興味深い系路をもつ生物のみを取り上げていく事にする。