
だが、話は「歴史生物学 TCA回路への異論 8」で終わらすことはできない。こんなブログを読んでいる人であれば当然ご存知だと思うが、KEGG(Kyoto Encyclopedia of Genes and Genomes)というデータベースは、膨大な情報を集積しているだけでなく、使いやすい形で提供しているサイトである。(近頃アップグレードがなされ私にとっては幾分使い難くなった。でも不平は言えない。)個人ユーザーとして、寝る前のかなりな時間をこのサイトで遊んでいる。これほどの情報にアクセスできるなど、若い頃には想像できなかった。サイトを構築された金久實教授とそのスタッフに、どれほど感謝してもしすぎることはないと思っている。そんな私が、真核生物(動物: 85種、植物: 38種、Fungi: 72種、Protist: 41種)、原核生物(真性細菌: 2594種、古細菌:165種)の計2995種の生物について、どのような TCA 回路を持つかを調べてみた。最終的に一月ほどかかったのだが、視力が落ちて、肩が凝った。マッサージに行くはめになっただけでなく、自らの無知を再確認させられるような被虐的楽しみの毎日であった。
原核生物に我々のものとは異なるTCA回路を持つ生物がいることは知ってはいた。だが、真核生物の中にも、そうした生物がいたのである。アピコンプレックス門に属する原虫類、マラリア原虫に代表されるグループの TCA 回路は左回りの系で、2-ケトグルタル酸までで止まっている。もっともこのグループは寄生性の原虫であるから、系が不完全でも良いではないかという議論が成立するとして無視するのは可能だろう。だが、そういうわけにも行かないようだ。例えばXenopus laevis(アフリカツメガエル)、実験動物として広く使われ切り刻まれている気の毒なカエルだが、このカエルに於いてはフマル酸とリンゴ酸の間が切れている。Micromonas sp. RCC299は海洋性ピコプランクトンとして知られている緑藻の仲間だが、この藻に於いてはクエン酸から2-ケトグルタル酸をつなぐ酵素が存在しない。珪藻の中にも同じ欠損を持つ種が存在する。ということから、少し丹念に調べてみることにした。
真性細菌、いわゆるバクテリアや古細菌を片っ端からみていくと、回路になっていないTCA回路(この表現は矛盾している)を持つ種は珍しいものではなく、かなりな割合の種が存在している。最初はそれほど多数にはならないと楽観していたのだが、余りの多さに驚いてしまった。この部分をきちんと割合で示す事ができればいいのだが、さすがに再度見直す気分にはなれない。ただ、TCA 回路の空白部分は、幾つかのパターンに分けられそうである。同時に、当然ではあるが寄生性の生物においては不完全な回路を持つものが多いようだ。
前置きはこれくらいにして、幾つかの欠損パターンを持つTCA(非)回路を示してみよう。図3-23にPseudomonas aeruginosaの持つTCA(非)回路を示す。
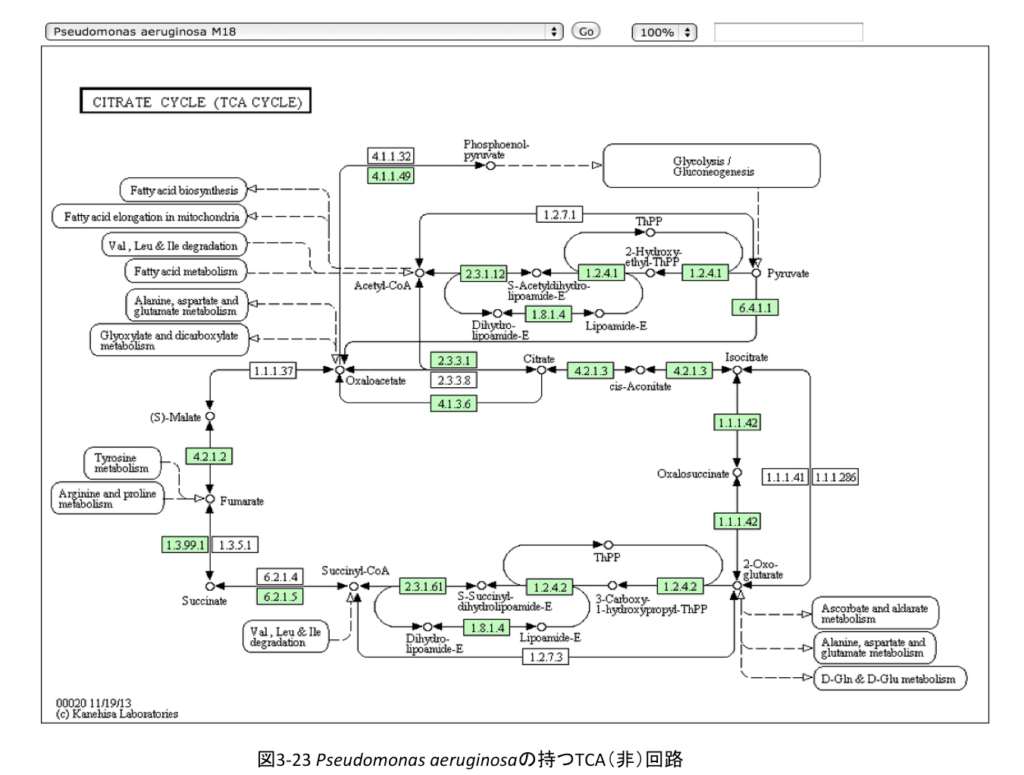
こうしてみるとリンゴ酸とオギザロ酢酸の間で切れているように見えるが、このタイプの系路を持つ生物には、ピルビン酸とリンゴ酸をつなぐ系が存在するため、系のメンバーとしてピルビン酸を加えれば回路が成立する。Pseudomonas の中では、Pseudomonas mendocina、Pseudomonas syringae、 Pseudomonas stutzeriがこの系を持つだけでなく、Staphylococcus aureusやDesulfosporosinus orientis、Marinobacter aquaeolei、Thiomicrospira crunogena、Allochromatium vinosum、Marinomonas mediterranea、Neisseria meningitidisなどかなりな数の微生物がこの系路を持つ。従って、このグループは修飾された TCA 回路を持つものと見て良いだろう。但し、それぞれの生物が持つ回路がどちらを向いて機能しているかについては確認していない。好気性の微生物においては右回りに機能していると予想しているが、この点は今後の課題になるだろう。
面白いのは図3-24に示すMethanothermococcus やMethanocaldococcus属細菌の持つTCA(半)回路である。
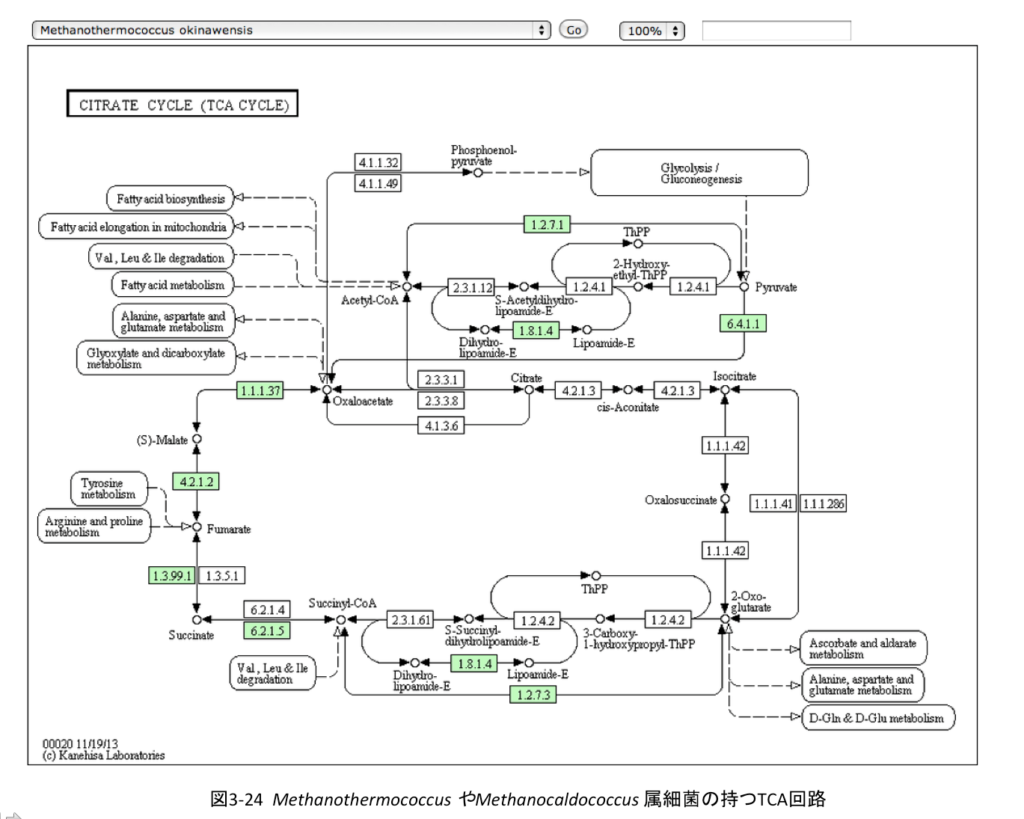
この古細菌において、系は左回りに駆動しオギザロ酢酸からα-ケトグルタル酸に到達する。そして右半分は存在しない。この左半回路を持つ生物は古細菌だけではない。真性細菌であるChlamydia、Chlamydophil、Bifidobacterium、Desulfotomaculum、Prevotella属細菌も、同じようにオギザロ酢酸からα-ケトグルタル酸までの左回りの代謝を行う。左回りに半分しか存在しないのに回路という表現は不適切だとは思い、左半回路という表現を用いた。
一方、図3-25に示すように、TCA回路の左半分を欠いている右回りの系を持つ生物も種々存在する。

Eubacterium、Roseburia、Coprococcus、 Ruminococcus 属細菌などである。彼らはオギザロ酢酸から右周りに系を駆動し、2-ケトグルタル酸まで代謝を行う。アセチルCoAの生合成系がないではないかと思われる方がいると思うが、これらの生物においては Acetyl-CoA:formate C-acetyltransferaseという酵素が、ピルビン酸とアセチルCoAの間をつないでいる。Synechococcus、Brachyspira、Prochlorococcus 属細菌もほぼ同じ右半回路系持つ。
これら以外に、上記の系路が1段階あるいは2段階延伸した系を持つ細菌、脈絡を考えにくいようなランダムな系を持つ細菌、TCA回路をかけらも持たない細菌などもいる。脈絡を考えにくいようなランダムな系を持つ細菌、TCA回路をかけらも持たない細菌類は、寄生あるいは共生という生活様式を持つものがほとんどを占め、ホストあるいは共生の相手にこの部分を依存していると思われる生物群である。(共生と寄生を明確に区別するのは極めて難しいが、ここでは常識的な用語として使っている。)
上に述べた現実を基盤としてTCA回路の意義を考える場合、TCA回路はエネルギー(ATP)生産系などという説明はもはや意味をなさない。意義については、幾分強引な推論だが、いわゆる解糖系と還元的カルボン酸サイクルに加えて、図3-24と図3-25の系について考えれば良さそうだ。これらの4種の回路・非回路を持つそれぞれの生物群において、それぞれの系は同じ目的で駆動されていると考えて良いだろう。幾つかの異なる代謝系が同じ目的を持って駆動している場合、まず各集団に共通するものを探すのが鉄則である。この場合、4つの系で共通している化合物は 2-ケトグルタル酸である。この化合物、団塊の世代としては α-ケトグルタル酸と呼びたい化合物であり、文中でも α-ケトグルタル酸と書いてしまった部分があるようだ。
どうやら、解糖系と還元的カルボン酸サイクル、右回りと左回りの半回路において、共通する構成成分は α-ケトグルタル酸であると考えて良さそうだ。ではこの α-ケトグルタル酸、生物の中でどのような意義を持つのか。多くの方がご存知の通り、α-ケトグルタル酸からはアミノ酸代謝が出発する。α-ケトグルタル酸のα-位のカルボニル基にアミノ基転移が起こりL-グルタミン酸、L-グルタミン酸の γ 位のカルボキシル基がアミド化されてL-グルタミンがつくられた後、これらが種々の α-ケト酸にアミノ基を転移して、多様なアミノ酸類の生合成が起こる。アミノ酸生合成は、生物がタンパク質をつくる前段階の欠くべからざる反応であり、「その出発物質であるL-グルタミンとL-グルタミンの原料である α-ケトグルタル酸を供給するのがTCA(非)回路の意義である」とするのが適切な判断だと思うが、諸氏の考えはどうだろう。
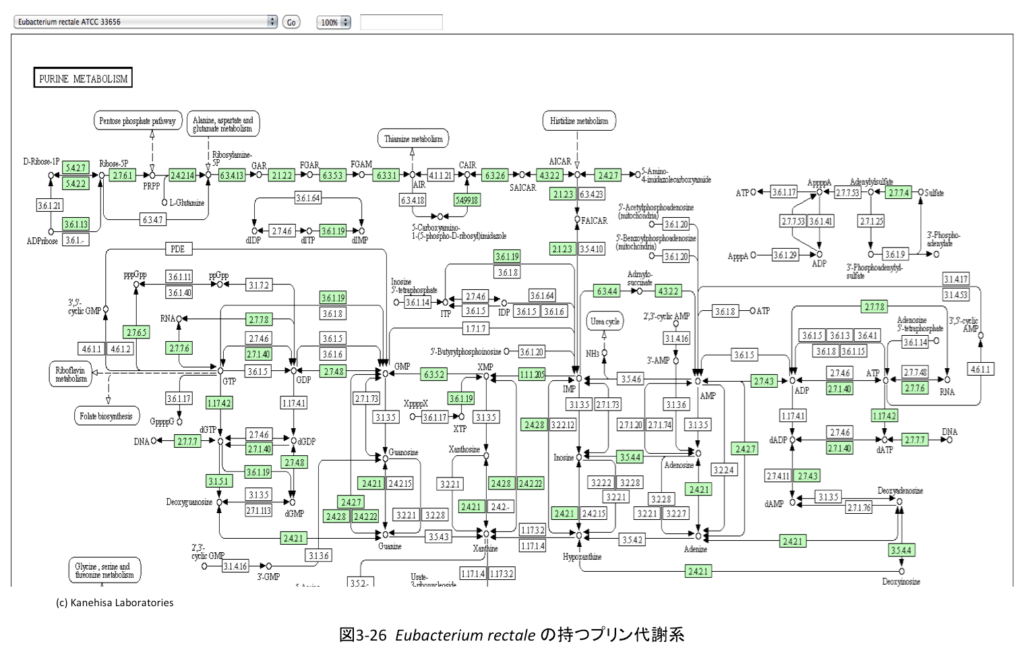
重要な補足である。α-ケトグルタル酸から誘導されたL-グルタミン酸とL-グルタミンの存在意義は、先に述べたアミノ酸生合成の出発点にあるだけではない。両化合物は、図3-26に示すようにプリン代謝につながりDNAとRNAの原料であるアデニン、グアニンの生合成につながるのみならず、ピルビン酸の脱炭酸段階で必要なチアミンリン酸生合成へと伸びている。
また両化合物は図3-27に示すとおりピリミジン生合成の原料ともなりながらDNAとRNAの原料であるチミン、ウラシル、シトシンへの生合成を可能にしている。

回路になっていないTCA回路を持つ多くの生物が存在している。酸化的リン酸化の系を持たないClostridium 属のような偏性嫌気性細菌にも、α-ケトグルタル酸で止まる右回りのTCA回路を持つものが存在する、もちろん左回りの系を持つ偏性嫌気性菌も存在する、右回りの系と左回りの系の共通代謝物はα-ケトグルタル酸である、α-ケトグルタル酸は種々のアミノ酸代謝のハブに位置する化合物である、α-ケトグルタル酸はプリン塩基とピリミジン塩基生合成の共通の原料である、という事実を重ね合わせてこの系の意義を推測するとすれば、第一義的にはα-ケトグルタル酸の供給という側面を見なければならない。エネルギー獲得系という意義付けは、酸化的リン酸化の能力を持つPseudomonas denitrificansのような好気的微生物や、Pseudomonas denitrificansの末裔ともいわれるミトコンドリアを持つ生物において成立するにすぎないだろう。但し、これらの生物においても、α-ケトグルタル酸の供給という系の意義は不変である。
そうすると、TCA回路・還元的カルボン酸サイクル・不完全なTCA回路に通底する根源的意義は、α-ケトグルタル酸の供給に求めざるを得ない。「生物の発生以来、右回りであろうと左回りであろうと、TCA 関連系路は α-ケトグルタル酸の供給を続けてきた。その後、酸化的リン酸化能力を獲得した生物が、系を右向きに回すことで生成するNADH2やFADH2を酸化し、多量のATP生産能力を獲得した。しかし、これらの生物においてもα-ケトグルタル酸の供給の意義は継続している」と記述すべきではないだろうか。
ギリシャ神話においてカイロスは両足に翼を持つ形で表されるが、私の想像(妄想)にも大きな翼が付いているようだ。時として、勇み足となる場合もあるだろう。しかし、結果を見た後で理屈をこねる経済学者のようにはなりたくない。夕暮れに飛び立つミネルバのフクロウにはなりたくないのである。そう思ってここまで来たのだが、1つ間違うとイカロスになる運命が待っているかもしれない。まあそれでも良い、クロノス時間に従っていても楽しくない。私の妄想はカイロスの時間に相性が良さそうだ。
歴史生物学 TCA回路への異論 完
しばらく、PowerPoint が昔つくったファイルを開けなくなっていた。古いPCを使っているため、色々と不都合が起こり始めている。新しいマシンを入れようかなと思わないわけではないのだが、マシン代に加えて、日本語入力ソフト、年賀状作成ソフト、ChemDrawあるいはChemOfficeなど、新たに購おうとすれば40万円、あるいはそれ以上の出費を覚悟せざるを得ない。一寸以上に躊躇する。身の程を考え、生活を質素なものにして、想像と妄想と創造の世界を豊かにしていく方が実りあるものになりそうな気がしている。