
以前に書いて公開していたはずのものだが、残っていないようなので再度公開します。4月1日の公開分ですので一寸怪しいかもしれないな。
昔といってもそう遠い昔ではない。植物は顕花植物と隠花植物に分けられていた。この分類はいまではなくなったらしく、花をつける植物は裸子植物と被子植物を含め種子植物として分類されるそうである。とはいえ、裸子植物の胞子葉をみてこれを花だと感じるヒトは、少なからず、いや過剰に植物分類学に毒されたヒトであろう。

我々が普通に花らしく感じる花は被子植物の花である。そして、我々が美しいと感じる花を咲かせる植物は、確かに多数存在する。ではなぜ、花は美しく進化したか。この疑問は果てしなく深い。ある花を、ヒトが美しいと感じるのは何故なのかと考え始めると、花の話に入る前に美とは何かというヒトの認識論に陥ってしまう。科学哲学を志そうとは思ったものの、本物の哲学者になる決心はまだついていないので、そこまで遡るのは止めることにしよう。
とりあえず、花はヒトにとって美しく感じられる色と形へと進化してきたという、多くの例外を含む言説があることだけは認めておこう。(スマトラオオコンニャク、ラフレシア、ザゼンソウ或いはマムシ草などの花を見て、あなたが美しい色と形であると感じるかどうかは分からない)。

では、花はなぜ美しいのか、あるいは美しくなる方向で進化したのか?この疑問に対して、世の中には虫によって花粉を運んでもらう虫媒花の話へとさり気なく誘い(いざない)、虫と花との共進化へと話を絞り込む先生・研究者などが極めて多い。某教科書会社の教科書によると「花がきれいなのはなぜ?」という単元に、「花粉を昆虫に運んでもらって受粉する花を、虫媒花 といいます。虫媒花は、虫をよぶための大きく色あざやかな花びらとあまい香り、虫たちへの報酬としておいしい蜜や花粉の一部を用意しています。虫媒花は、風まかせの風媒花より効率よく花粉が運ばれ、受粉の確率が高くなり、確実に種子をつくることができます。」と書いてある。
この説明において、美しい・きれいという表現を使わずに「大きく色あざやかな」としたのは、ムシにとって美しいという概念は使えるのかという質問から逃れるためのずる賢い選択であると受け取れない事もない。しかしながら、大きく色あざやかと書いても何も変わらない。大きく色あざやかに感じる主体はムシであろうかヒトであろうか。そして次に使われている「あまい香り」も「美味しい蜜や花粉の一部」というフレーズも、感じる主体を曖昧にすることで成立している。
著者はホモサピエンスに属する1個体として、蜜は場合によっては美味しいことを認めるが、主食にするようなものではない。10年ほど前に血糖値が高くなり、それ以来食べていないような気がする。花粉については美味しいかどうか判断できるほどまとまった量を食べたことがない。健康食品的扱いで売られている花粉があるようだが、大多数の人も食べたことはなかろうと推測する。そうすると、さきの虫媒花の定義は植物がムシたちの嗜好を了解した上で、「報酬」として蜜と花粉を用意したという話になる。しかし、花で分泌する蜜だけ吸って花粉を運ばない虫も多数存在する。さらに、ムシが人にとって甘い香りに引かれることがあるのを認めないわけではないが、同時に人が忌み嫌う臭いに引かれるムシも多数存在する。
私見だが、上記の説明は、地球上での虫媒するムシの出現と虫媒花の出現はどちらが早いのか、植物による蜜の分泌と蜜を好むムシの出現はどちらが先なのか、ムシは風媒花の花粉を食べても良いではないか、花外蜜腺の存在をどう説明するか、風媒花の植物でも十分に繁栄してるのは何故か、イネ科植物は虫媒花から風媒花へと進化した事実をどう説明するか、などなど基礎的な議論を無視した所に成立する何とも理解不能な詭弁である。子供相手とはいえ教科書である。いい加減に書いたとは思わないが、これら教科書の執筆者・監修者がかなり高名な方々である事を考えると、ちょっと杜撰、いや無責任すぎるのではないだろうか。さらに、一般人の検索を意識して書いてあるウィキペディアにおいても、「虫を誘引するために美しく目立つ姿や強い香りを放つものが多い。また、蜜を出すのも虫を誘引するための適応と考えられる。」と書かれている。
どうやら、ムシとヒトは審美眼と味覚、さらに嗅覚においていくつかの例外を除けば似た判断をしているというのが世の常識であるらしい。ムシとヒト、確かに遺伝子レベルではかなりな相同性を持つ。ムシで足の発生を誘導する遺伝子とヒトの手足を誘導するマスター遺伝子は相同であるし、ムシの目を誘導する遺伝子と人の目を誘導するマスター遺伝子も相同である。(これはこれで、生物学における相似と相同の概念に影響を与える悩ましい現象である)そしてDNAからRNAを通ってタンパク質を作る情報伝達メカニズムもほぼ同じである。
しかしながら、一部のムシは我々の目では見ることのできない紫外線まで見ることができるようだ。ヒトの目ではモンシロチョウの雌雄を迅速に見分けることは難しいが、モンシロチョウの雄は我々にとっては紫外領域にあたる光の反射特性によって、瞬時に雌雄の判別をしているという。大まかに言えば、昆虫の可視域はヒトの可視域に比べてでは、100 nm程短波長側にずれているそうだ。我々にとって真紅のバラは、昆虫にとっては漆黒のバラであるかも知れない。そのあたりはムシに聞いてみないとよく分からない。とにかく、見ることのできる波長範囲が異なる生き物に、審美眼が同じといわれても納得できないとモンシロチョウは言うかもしれない。
議論になっているいろいろな事項の、地球上に現れた時期についての議論は一旦横に置くとして、花、特に虫媒花の説明に関しては、あまりにも擬人化が過ぎるのではなかろうか。私だって擬人化できないわけではない。例えば、虫がつくという表現があるが、このムシとは読者が容易に妄想できるムシである。花にはもちろん“花”という暗喩がある。このムシ達が、甘い香りと美しく色鮮やかな花に惹かれて集まるのである。花は甘い蜜と花粉のパンをムシたちへの報酬として用意している。ときには有毒な花もある、ムシを捕まえて食べてしまう花もあるとすれば、いやいや花もムシもネオン煌めく夜の巷に棲息しているようだ。昔に戻って、植物採集を再開したい気分になってしまうが、食虫植物の捕獲対象になる可能性の方が高そうだ。歴史生物学というテーマで生真面目にライフワークを展開しているつもりだが、お花畑=夜の街論になってしまったようだ。
原点に戻ろう。生物的現象を歴史的視座から見るという原点に立ち戻ったとき、複数の現象はどのような順番で出現したのかを問わねばならない。コケ植物やシダ植物を観れば植物が初めて花という器官をつくったとき、それが風媒花、いや多分水媒花であったことは間違いないであろう。しかし、水媒あるいは風媒花から虫媒花への進化が起こるに際して、花が出し始めたかもしれない香りを感じる能力を持ち、かつその発生源に蜜と花粉があることを知っていた昆虫がいたのであろうか。前適応?もちろんそれに類した能力を虫が持っていたであろうことは推測できる。しかし、そうした性癖を持つ昆虫を引き寄せ花粉の運搬をさせようとする意図をもって、植物が香りの成分を作り花弁に色を持たせ、少々食べられても良い量の花粉を造ったとする説明は、わかりやすい嘘だと思う。前適応と言う万能の言葉で安易に議論を終わらせる、それは嘘つきの知的怠慢を意味している。
子供の頃に怪傑黒頭巾(だったと思う?紫だったかな)というテレビ番組が放送されていた。捕方に追われ屋根の上を逃げていた黒頭巾が追い詰められてくると、必ずそこの屋根の下にアオという名の馬が待っていたように記憶している。あんな馬が欲しいと思ったものだが、それはドラマのなかの約束事である。あれは馬が待っていたのだろうか、それとも馬のいるところに逃げて行ったのだろうか?
植物が花粉や蜜をつくったからといって、それを運ぶ昆虫が都合良く待っているわけはあるまい。花が大きく色鮮やかでかつ強い香りを持ち、蜜をも分泌するようになったのは、花を咲かせる植物の側にそれなりの事情があったに違いない。ムシが寄ってきたのは、花ができた後の話である。2つの生物間のそれぞれの事情がどこかで交錯し、共進化と呼ばれる現象が開始されたことを否定する気はないが、植物側が昆虫が起こすであろう行動を予測して始まった事ではないだろう。もちろん、昆虫側も植物はこんな物質群を造るだろうと予測して、待っていたとは思えない。少し前までは昆虫の行動を、本能と学習という分かったような分からないような概念を使って安易に説明していたが、それら起源を厳密に問われると自己撞着に陥ってしまう。現在の時点で昆虫側の話は私には出来ない。昆虫を抜きにして、植物が何故色鮮やかな花を創ってきたのかを考えてみたい。
そこで花を創った植物側の事情とは何か。花の色とは何かという疑問である?花の色を決めているだけでなく、植物体内に広く分布する色素は生合成されるルートによって、三つのグループに分類できる。一つはカロテノイド(前節で述べたテルペノイドに属する化合物群でカロテン、キサントフィルなど植物だけではなく、動物、微生物などにも広く分布する色素群)、もう一つはフラボノイド(フラボン、フラボノン、フラバン、イソフラボン、アントシアニジン、カルコンなどを含むC6-C3-C6ユニットを持つ植物に分布する色素群)、そしていま一つは、分布は狭く砂糖ダイコン、ヨウシュヤマゴボウ、ホウレンソウ、ウチワサボテンなどに含まれるベタレインと呼ばれる3種の色素群である。緑色の花もないことはないのだから、この3種にクロロフィルを加えるべきかもしれないが、クロロフィルは一般的には花の色素として認識されていない。クロロフィルについては3種の色素の話が終わったところで少し触れることにする。そこで、主要な3種の色素について少し述べておきたい。
カロテノイド
カロテノイド(carotenoid)、語源はcarrot(ニンジン)であり、carrotの語源はギリシャ語のkaroton、ラテン語のcarotaに由来するそうだ。カロテノイドは前節で述べたように、多数の共役した二重結合を含む化合物群で、テルペノイドの一群として分類される。主として赤色、橙色、黄色を示す色素群であり、ガーベラ、キク、マリーゴールド、キンセンカ、ガザニアなどの黄色から橙色の花の色素である。自然界では、高等植物だけでなく蘚苔類、藻類、あるいは酵母、バクテリアなども生合成能力を持ち、地球上では年間に約1億トンが生産されているという。花の色素として機能するカロテノイドには、α-カロテン、β-カロテン、リコペンなどを含むC40カロテン類と、それらの酸素添加反応生成物であるルテイン、ゼアキサンチン、ビオラキサンチンなどを含むC40キサントフィル類などが存在する。
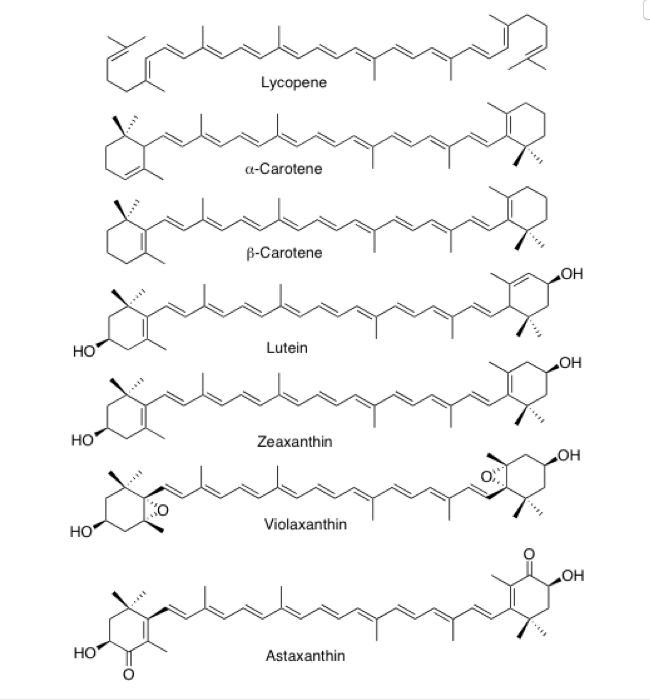
実に味気ない叙述である。間違ってはいないかもしれないが、面白くも何ともない。書いた本人がそう思うのだから、読む方はもっと退屈に感じ、もう睡魔に懐柔されているのではと勘ぐりたくなる程である。実はこの先に、45や50個の炭素原子を含むカロテノイドがあるとか、キノコにも含まれる、サケ・マスの肉の色あるいはエビ・カニを煮たときに赤くなる甲羅の色の成分もアスタキサンチンというカロテノイドのせいだ、フラミンゴの婚姻色はβカロテンが原因だなどと並べることは可能だが、それではこの文章は睡眠薬にしかならないだろう。
さりとて、現世の健康第一主義、健康のためなら命もいらぬと言う風潮に靡いて、β-カロテンは健康によい、ビタミンAの原料になる、ルテインは目によい、リコペンはに抗酸化作用があるなどと、サプリメント業界に媚びを売るつもりもさらさらない。実はここからどう話をつなごうかと、書きながら考えている。面接で、思わぬ質問を受けたとき、「思いがけない質問で少し困っています。とはいえ、その問題には考えるべきいくつかの側面があると思います。その中で・・・・・」などと意味のない言葉を連ねながら、答えるべきことを必死に考えている状態と似ているようだ。
先に述べたようにアブシジン酸生合成の中間に位置するこの β-カロテンは、以前はアンテナ色素として植物の効率的な光の吸収と伝達で働くのが主な機能と考えられていた。しかし近年では、光合成に伴って生成する活性酸素の生成を防ぐとともに、生成してしまった有害な活性酸素から植物自身を保護する作用のほうがより重要である事が明らかになってきた。
そこで光合成だが、光合成反応は明反応と暗反応に大別される。と心置きなく書けたのは20世紀の終わり頃までだった。いまはどう呼ぶか、明反応の部分は光化学系とこれに連なる電子伝達系、暗反応の部分は炭酸固定・炭酸同化あるいは炭素固定・炭素同化などが使われているようだ。理由は簡単で、明反応の中に光が関与しない部分があるから光反応は適当ではない、暗反応においては、暗いところでは明反応からのエネルギーと還元力の供給がないだけでなく、暗反応の主要な部分を担うカルビンベンソン回路に含まれる酵素の一部が光による制御を受けており、暗反応は暗所では作動しない。従って、暗反応という名称は適切でないということであるようだ。いやいや、ややこしいことになってきた。電子伝達系には、光合成と関係なく機能する場合があるから不適切だ、炭素固定にもTCA回路を逆向きに回す還元的カルボン酸回路やメタン発酵なども含むから不適切だなどと続けていったら、何処まで分ければいいのだろうか。
炭素固定に関与する酵素群が触媒するいわゆる暗反応は、素直な化学反応の連なりとして有機化学的に理解するのはさほど難しくない。ところが光化学反応は、私にとってと限定した方が適切かもしれないが非常に難しい。この反応において、電子の伝達を定量的に記述した本はほとんどないのではなかろうか。これは植物生理学といういくぶん生物学よりの学問を専攻した方々が、電子伝達を電子論的に記述することに不慣れであったこと、電子伝達系で働く化合物群の分子量がかなり大きく、図として描くことが困難であること、さらにこれら化合物群が共役した多数の二重結合を持つため反応に関与する電子をどこに描けばいいのか判断しにくい、などに起因していると思われる。
さらに、明反応における電子の励起・伝達の部分は、いわゆる有機電子論の域を超えて量子化学の領域にある。電子が何処にあるかだけではなく、電子のスピンをも考慮して捉えないといけない。此処にいたって、筆者の理解は概念的なものになってしまう。一重項酸素と三重項酸素の区別はつくが、励起一重項クロロフィルと三重項クロロフィルの違いをどう描くかとなると、??である。HOMOとLUMOの世界である。その程度の理解で以下の文を書くため、間違いがあるかもしれないし、表現に不適切な部分があるかもしれないことをお断りしておく。(教育的指導をしていただければ、なお有り難い)
さて、シアノバクテリアだけでなく全ての植物では、光合成中心にあるクロロフィルは光エネルギーを吸収して励起状態となった後、励起された電子を放出する。正常であれば、この励起された電子は電子伝達系を流れながら、ATPの生産あるいはNADPH++H+の生産に用いられる。と安易に書いてしまったが、これでは正しくないか。光化学系Ⅰと光化学系Ⅱに分けて考えないといけない。ここは後で書き直すとして、一部の励起一重項クロロフィルは三重項クロロフィルと呼ばれる活性化された状態になる。この三重項クロロフィルは三重項酸素と衝突すると、電子とエネルギーの交換を起こし、クロロフィルが基底状態に戻ると同時に、三重項酸素が有害な一重項酸素に遷移する。β-カロテンは、この三重項クロロフィルのほとんどを元のクロロフィルに戻すと同時に、生成した一重項酸素とも反応して三重項状態の酸素に戻す役割を果たしている。β−カロテンが受け取ったエネルギーは熱あるいは燐光として廃棄される。あれ、私は何を書いているのだろうか.論旨と関係のないことを書いているようだが、普段から良い光合成関連の本がないという不満が噴き出したようだ。
少し本論に近づこう。植物とは、10億年ほど前に真核細胞の一種がシアノバクテリアと共生することで生じた新たな生物である。そして葉緑体(クロロプラスト)は、このとき取り込まれたシアノバクテリアの末裔である。ところでこのシアノバクテリアが酸素発生型光合成能力を獲得したのは35億年ほどまえであり、その光合成中心にはすでにβ-カロテンが存在したのは間違いない。(この年代推定も必ずしも正しいというわけではない。シアノバクテリアの出現時期に関しては、27億年前、32億年前、35億年前、37億年前と、いろいろな説があり確定しているわけではない。ただ、報告が新しくなるにつれて、より古い時代へと移行しているような感はある。35億年の値を採用する)この事実とβ-カロテンの生合成に酸素は不要であることを考え合わせると、地球上でのβ-カロテンの歴史が35億年以上になることは自明であろう。
つまりβ-カロチンは、35億年ほど前に始まったシアノバクテリアによる酸素発生型の光合成反応において、三重項クロロフィルを消去するとともに、残った三重項クロロフィルから生成してくる一重項酸素を、比較的安定な三重項酸素に変換し続けてきたと言える。この2つの役割を果たす際に、β-カロテンはエネルギーを受け取って三重項状態に励起されるのだが、受け取ったエネルギーは熱あるいは燐光として捨てられるためβ-カロテン分子自体に化学変化は起こらない。ところが、20億年ほど前に出現した酸素添加酵素(オキシゲナーゼ) が βカロテンを基質として認識したために、β-カロテンはクリプトキサンチン、ゼアキサンチンを通ってキサントフィルと呼ばれる化合物群へと酸化されることになった。(更にだが、最初に出現したシアノバクテリアが持っていたRubiscoにオキシゲナーゼ活性があったかどうかの問題である。もし持っていたとすれば、オキシゲナーゼのレゾンデートルも書き換えられるかもしれない。)
ここにおいてβ-カロテンは、過剰なエネルギー廃棄の通路となるだけでなく、自らの分子中に酸素を取り込んで、酸素分子自体を消去する役割をも担うことになった。さて、β-カロテンからほんの1段階酸化が進んだに過ぎないクリプトキサンチンは、この視座から見ると最大に見積もっても20億年の歴史しかない。つまり、β-カロテンからクリプトキサンチンへの1段階の代謝の伸長に、少なくとも15億年という時間が必要であったことを示している。この後クリプトキサンチンは、さらに5段階の酸化を受けてアブシジン酸に導かれるわけだが、そう考えるとアブシジン酸が若い分子であることは自明のことであろう。
シアノバクテリアは鉄硫黄型反応中心をもつ光化学系Ⅰとキノン型反応中心を持つ光化学系Ⅱを持つ。前者は緑色硫黄細菌に由来し、後者は緑色糸状菌あるいは紅色光合成細菌に由来する。緑色硫黄細菌のすべてがβ-カロテンまでの生合成系を持つことを考慮すれば、カロテノイド生合成系は35億年よりもかなり前にβ-カロテンまでの系が完成していたことを意味する。同時に、β-カロテンの段階で、15億年以上足踏みをしていたのである。アブシジン酸が若い分子であると言うことが、今後の話の中で大きな意味を持ってくるので、是非頭の隅に残しておいて欲しい。
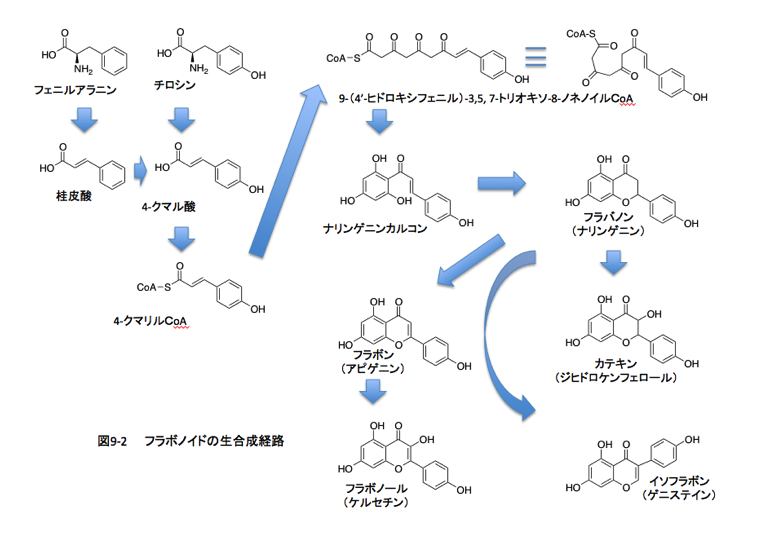
本論に戻る。次はフラボノイドである。フラボノイドとは黄色、橙色、青色までの広い色調を示すC6-C3-C6のユニットを持つ化合物群で、フラバノン、フラバン、フラボン、フラボノール、イソフラボン、アントシアニジン、カルコン、オーロンなどを含み、配糖体まで考慮すると7000種を超す化合物で構成される極めて多様な物質群である。
フラボノイドについて
次に簡略化したフラボノイドの生合成系の図を示す。ただ、この図が全てを網羅してないことに注意して欲しい。桂皮酸からシンナモイルCoAを通ってクマロイルCoAになる経路を省いているし、コーヒー酸からの流れも省いている。ダイゼイン生合成系も省いている。何故そんなに省くのかって?話の大筋に殆ど影響がないと考えたからである。カーネーション、キク、バラ、ビオラなどの花色の成分となっているとともに、ほとんど色のないフラボンやフラボノールが白い花の花弁に存在している。ではなぜ植物はフラボノイドを作るのか。またおなじみの疑問だが、答えは少し待って欲しい。フラボノイドの生合成を含めた少し詳しい話は後回しにして、まず常識的な予備知識を提供しよう。(ご存じの方は、ごめんなさい。)
フラボノイドは太陽光に含まれている有害な紫外線から植物を守るといわれている。太陽の表面温度は6,000℃、強烈な紫外線を周囲に空間に照射しているわけだが、先ほど述べたように6億年ほど前の酸素濃度の急上昇に伴うオゾン層の成立によって、この紫外線の地表への到達量が激減した。これで生物の、上陸への準備が整ったといえる。しかし、UV-Cと呼ばれる短波長のエネルギーの高い紫外線はオゾン層でほぼ完全に遮られるにしても、UV-Bと呼ばれる280-315 nmの紫外線の約0.5%と、UV-Aと呼ばれる315-380 nmの紫外線の5%あまりは、地表まで透過してくる。UV-BはDNAに光反応を誘発し隣接するピリミジン塩基間で2量体の形成を引き起こすし、UV-Aはタンパク質の変性を誘発する。従って、オゾン層が成立したとはいえ、生物は上陸に際してこの紫外線への防御機構を持つ必要があった。
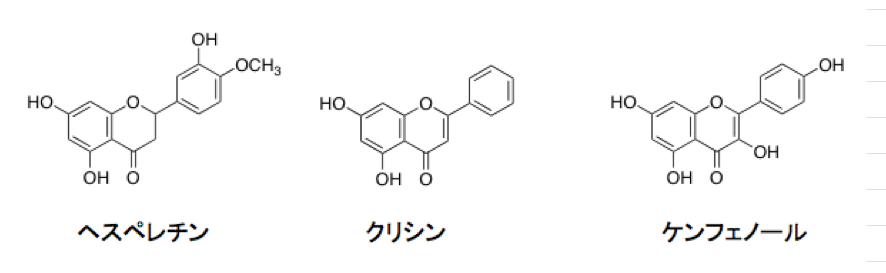
植物は葉の表皮細胞で紫外線をよく吸収するフラボノイド(特にUV-A, UV-Bをよく吸収するフラボン、フラボノ―ル)を生合成する。そして確かに、フラボノイド類は紫外線障害から植物体を守っているようにみえる。フラボノイドを合成できない変異種は、紫外線に弱く太陽光の下では生育が抑えられることも知られている。色のないフラボノイド、例えばヘスペレチン、クリシン、ケンフェノールなど可視部に吸収を持たない化合物群でも、有害な紫外部に強い吸収帯を持ち、紫外線に対する防御に役立っているのは間違いないであろう。
透明な花がある。サンカヨウ(山荷葉)というメギ科の多年草で本州中部以北に分布する。白い花だが水に濡れると花弁が透明になる。それは我々が見た場合であって、UV域では透明では無いと考えているのだが、北日本のどなたか、透明になった花弁のUV吸収スペクトルを取ってはいただけないだろうか。
またもや余談だが、フラボノイドの基本となる骨格上にはいくつかの水酸基が存在する。これらの水酸基の中で5位、7位の水酸基は、ポリケチド鎖が閉環、芳香化するときにケトーエノール互変異性体のエノール体として現れたもので、酸素分子が関与する酸化生産物ではない。同様に、4‘位の水酸基はアミノ酸であるチロシンの水酸基に由来しており、オキシゲナーゼが関与する水酸基ではない。
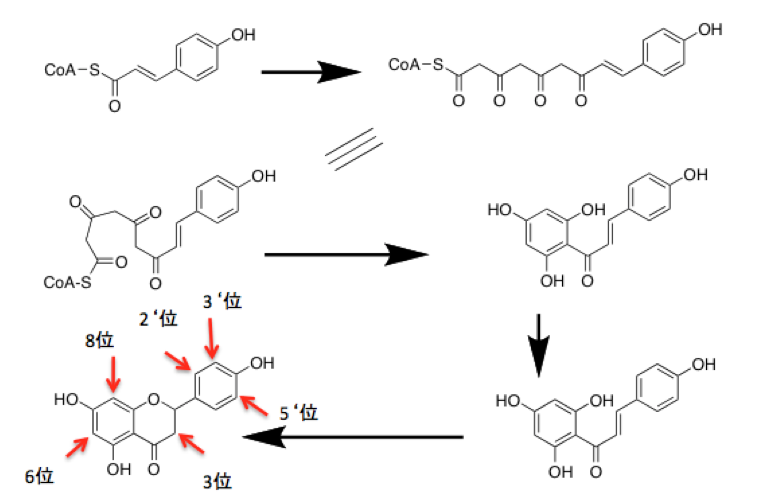
従って、4’, 5, 7-ヒドロキシフラボン(通称: ナリンゲニン)は、嫌気的に生産可能な代謝物である。ところが、自然界には、2‘位、3‘位、5’位あるいは3位、6位、8位などに水酸基を持つフラボノイドが存在する。これらの水酸基は、先に述べた水酸基とは異なり分子状酸素の存在下にオキシゲナーゼの作用によって導入された水酸基である。フラボノイド分子から水酸基を除く酵素の報告はないことを考えると、フラボノイド分子を酸化程度に従って並べれば、それはフラボノイド分子を進化の順序に従って並べたものになることを意味している。
サントリー(株)が作出した青いバラ(私には赤紫色にしか思えない)は、パンジーの持つ3’位と5’位に酸化的に水酸基を導入するフラボノイド3′,5′-ヒドロキシラーゼ遺伝子をバラに入れ、バラ花弁でデルフィニジンの生合成を可能にすることで成功した。これは、酸素を消去する代謝過程を一つ付け加えたことを意味する。従って、この青いバラは、元のバラに比べ酸素に対する抵抗性がちょっぴり上がっているに違いない。
ベタレイン色素
ベタレイン系色素は植物での分布は狭く、主とし、アントシアニンを生合成できないナデシコ目植物に分布する。我々になじみ深いのは、マツバボタン、ケイトウ、オシロイバナ、それからホウレンソウ、サボテン、ブーゲンビリア、変わったところでは菌類に属するベニテングタケの真っ赤な色の成分などである。
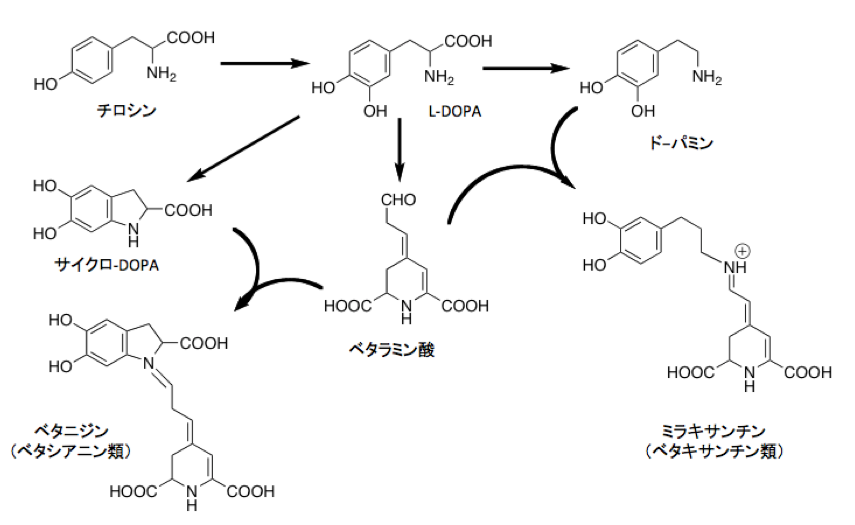
上図に示すようにこのベタレイン系色素は、植物体内でチロシンから生合成される。まずチロシンにチロシナーゼと呼ばれる水酸化酵素が働きL-DOPAとなった後、4,5-DOPA dioxygenase extradiolの作用を受けてベタラミン酸が生成する。このベタラミン酸がドーパミンから誘導されるcyclo-DOPA類と反応して赤から紫の色調を持つベタシアニン類が生成する。一方、ベタラミン酸がプロリンのようなアミノ酸あるいは生体アミン類と反応すると、黄色からオレンジ色の色調を持つベタキサンチン類が生成する。ベタシアニン、ベタキサンチン類ともに、ベタラミン酸のアルデヒド基がベタラミン酸アミノ基あるいはイミノ基と縮合反応を起こして、共役によって安定化されているとはいえ酸化されやすいシッフ塩基あるいはイミニウムカチオン構造を持っている。
このベタレイン系色素、人の代謝と比較すると興味深い。人において、チロシンから生成するL-DOPA、L-DOPAの脱炭酸で生成するドーパミンは精神状態を興奮側へ導く化合物群で、L-DOPAはパーキンソン氏病の治療薬でもある。ドーパミンからはノルアドレナリン、アドレナリンの生合成系がつながり、交感神経を興奮させるホルモン類の原料となっている。この系統の化合物を少し修飾してやると覚醒剤として働くようになる。同時にドーパの酸化で生成するドーパキノンも非常に反応性が高く、人のメラノサイト細胞の中でドーパクロム、インドールキノンへと非酵素的に変化した後、酸化重合を繰り返して真性メラニンと成り、人の皮膚の色素となる。色の黒い人はここの代謝系に多くの物質が流れていることを意味するだろう。そういえば、色の黒い人に鬱ッポイ人は少ないように感じている。とはいえ、ナデシコ目植物の細胞内で、人の精神状態を左右し、皮膚の色を決めるような反応が動いているとはなかなか面白い。
さて、2017年に日本農芸化学会から出版された化学と生物に、「植物色素ベタレイン—分布,生合成および生理機能ー謎に包まれた多機能性植物色素」と題する紹介記事があった。近年までに得られた知見がよくまとめられており、読みやすい記事だとは思うものの、2つの点で物足りなさを感じる。1つは植物がなぜベタレイン系色素を作るのかという疑問を明確に意識することなく、創られた後で発現したベタレインの生理活性を基礎としてその存在意義が議論されているという点にある。ベタレインの持つ生理活性が、その生合成能を獲得した植物において有効に働いた事を否定するつもりはないが、そのことがベタレイン創製の理由にはならない。時間的な経緯を考えれば、創られることが先行し生理活性の獲得あるいは発現はその後に起こったことである。
いま一つの物足りなさは、著者の意識がチロシンを出発物質とし最終産物をベタレイン系色素とするベタレイン生合成系という常識に拘束され、その中でベタレインが活性酸素の毒性からの防御、花粉媒介者誘引のための視覚的シグナル、環境適応にかかわる生理学的な機能を議論している点にある。チロシンを出発物質とし最終産物をベタレイン系色素とするベタレイン生合成系という概念は、代謝の流れの中からベタレイン類に興味を持つ研究者達の極めて恣意的な分節に由来しているに過ぎず、植物の代謝系をそのように区切る必然性はない。例えばだが、イソペンテニルピロリン酸を出発物質とし、フィトエンまでをフィトエン生合成系とする、β-カロテンまでをβ-カロテン生合成系とする、ゼアキサンチンやビオラキサンチンまでをそれぞれの生合成系とする、アブシジン酸までをアブシジン酸生合成系とする分節は、研究者がどの化合物に興味を持っているかに支配されている。アブシジン酸の生合成系はイソペンテニルピロリン酸を出発物質とするには長すぎるとして、これをβ-カロテンまで繰り下げている場合もある。
ここから本論
花色を決めている色素類についての簡単な説明は終わった。そこで、本論に戻り花の色についての議論を始めよう。花の色が様々であるのは、「花に種々の色彩を持つ色素が存在するからだ」と一般には考えられている。しかし色素の存在は十分条件であり、色素があるからといってその色に見えるものではない。まず必要条件として、花の部分に葉緑素が存在しないことが求められる。(極少数の淡緑色の花も、葉緑素量が減っているものとして捉える) 通常、植物の緑色の部分にもいろいろな色素が存在しているが、葉緑素の存在量が際だって多いため緑色に見えている。例外的には、花色色素の存在しない白い花もあれば(白色色素があるという考え方も成立する)、水に濡れると花弁が透明になってしまう花もある。(http://nyanko.iza.ne.jp/blog/entry/42976/ 参照)つまり、植物体から葉緑素を除いた時点で、残りの色素が見えてくるのである。秋の紅葉は、ロマンチックなものでもメランコリックなものでもなく、葉緑素が分解された条件下に、フラボノイド類の分解産物であるアントシアン類の色が見えてきたすぎない。
さて花に存在する色素は、特定の波長の太陽光を吸収しながら、それ以外の波長の光を反射する。我々は反射された光を見ているわけである。赤い花に含まれる色素は、緑色、青色の光を吸収し、赤色光を反射する。反射された赤色光を見て、我々は赤い花だと感じるというわけである。植物の葉っぱが緑であることから、植物は光合成に緑の光を使っているように勘違いしている学生をよく見かけたが、植物は緑の光は利用せず、反射したり透過させたりして捨てているに過ぎない。光合成に使われる光は赤色の光と青色の光である。赤い光の下では、植物の葉っぱは真っ黒である。されば、美しいかどうかは問わないにしても花はなぜ色素を持つかという問いの裏側には、花はなぜ葉緑体を失ったのかという問いが存在すると考えた。とすれば、花はなぜ葉緑体を失ったのかという問いにまず答えなければならないだろう。
さて、花の定義を学問的に厳密に行おうとすると実に難しい。生殖に関連する器官は、いわゆる高等植物だけではなく蘚類、苔類にも存在する。まず、花という概念に何を含ませるかという定義以前の問題がある。常識的な範囲で考えても、被子植物の生殖器官とするのか、被子植物と裸子植物を含めた種子植物の生殖器官とするのか、あるいはシダ植物の胞子茎まで含めるのかによって、定義は大きく様相を変える。特殊な進化をした例外的なものを全て包含させようとすると、却っておかしくなってしまう。
私論だが大筋で間違わない程度の定義をするとすれば、茎の先端に雄しべ(葯と花糸)や雌しべ(柱頭、花柱、子房、胚珠)のような特殊化された器官を持ち、花弁や萼で包まれた種子植物の生殖器官とでもなるのだろうか。では、花の機能はと問われれば、生殖に伴い次世代を担う種子あるいは胞子の生産と考えて良いであろう。
この機能を基本に考えれば、蘚類や苔類の胞子体も同じ機能を持つ器官のカテゴリーに含めて良いだろう。面白いことに、蘚類や苔類の胞子体も多くの場合葉緑体を失っている場合が多い。誰もが知っているスギナの胞子体−ツクシに葉緑素はない。ゼニゴケは雌雄異株でそれぞれ葉緑体含量の少ない雌器托と雄器托を形成するが、その雌器托下に形成される胞子体は黄色であり葉緑素は含まれない。スギゴケも種類が多いとはいえ、胞子体の緑色はうすく褐色系統の物が多い。そうすると、花には何故葉緑体が存在しないかと言うという問いは、植物の生殖に関する器官には何故葉緑体が含まれないかと言う問いに置き換えられそうだ。
もちろん、この議論の進め方が幾分強引だとの批判はあると思う。だが議論を進めるに際して、無視すべき少数の例外と無視してはいけない少数の例外の峻別は避けて通れない。ではその峻別の正しさはどうすれば分かるか。歴史に委ねるしかないであろう。峻別という言葉を使うにしても、峻別するのは人である。人の行う峻別という営為は、通常主観による恣意性の発現に過ぎない。こうした諦観を持って、私は少数の緑色の花の存在をしばらく無視することにする。この部分を書いたのは平成13年の7月頃である。いま書くとすれば、排除と書いたかも知れない。流行りですから。
植物は細胞内に葉緑体を持つ。この酸素を発生する細胞小器官の存在故に、光合成を行っている植物細胞内の酸素濃度は非常に高い。また植物が光合成を行うに際して、受光したすべての光エネルギーが糖生産に使われるわけではない。強光下においては70%を超える光エネルギーが何らかの形で捨てられている。しかし、このエネルギー廃棄系から漏れ出すエネルギーが存在する。先にも述べたが、光化学系Ⅱにおいては励起されたクロロフィルが三重項酸素分子を一重項酸素分子へと励起する。この一重項酸素分子は反応性が高く、脂質の過酸化などを通して膜の破壊を引き起こす。光化学系Ⅰにおいては、励起クロロフィルが電荷分離を起こした後、電子伝達系に送られるはずの電子が酸素分子へと渡されると活性酸素の一種であるスーパーオキシドアニオンラジカルが、活性酸素防御系がうまく働いていない場合にはより反応性の高いヒドロキシラジカルも生成する。
一重項酸素、スーパーオキシドアニオンラジカルにSOD(スーパーオキシドディスムターゼ)と呼ばれる不均化酵素が働いて生成する過酸化水素、そしてヒドロキシラジカルなどは活性酸素と総称され、膜を破壊しタンパク質を変性させDNAを酸化する能力を持ち生命活動に悪影響を与える。これらが、ヒトを含む全ての生物の経年劣化の主要な原因であることが、近年よく知られるようになってきた。
酸素発生型光合成を行う植物において、高度に発達した活性酸素消去系が存在していることが求められる。つまり、植物細胞が葉緑体という光合成行う細胞小器官を持つと言うことは、CO2を固定して糖を生産するというメリットとがあると同時に、細胞が光を受けた時に毒性の高い活性酸素の生成が起こるというデメリットとの微妙な平衡の上に成立していると考えるべきである。こうした観点から見れば、減数分裂を伴う卵細胞と精細胞の作成と受精という生物にとって最も変異に弱い精妙な段階を遂行する場としての生殖細胞が、葉緑体を失うことは明らかに有利なことであったと思われる。
結果論ではあるが、葉緑体を失った生殖細胞を持つ植物の方が、有害な突然変異の影響を受けることなく子孫を残し続けてきたと理解して良いだろう。こう考えることによって、苔類や蘚類の胞子体、種子植物の花と呼ばれる器官が、葉緑体を失っている場合が多いことを体系的に説明できるのではないか。
さて、アメリカの心理学者ドン・クリフトンが発表した人間関係の理論に“バケツ理論”というのがあるそうだ。バケツの穴理論ではない。「人は皆、大きな心のバケツをかかえている」というものだ、このバケツに人からの称賛、肯定、認知、関心の水をためれば、その人は幸せになる。人から受ける非難、批評、悪口、叱責、無視等の否定はバケツに穴を開けるという。そして、バケツの中の水を失うと人は不幸になるという理論である。何となく異常に馬鹿馬鹿しく感じて殆どフォローしていないため、理論の正否についてというよりこれが理論であるかどうかさえ判断できない。水の入ったバケツなんて、持たされ立たされた記憶があるのみである。その場合、水は少しでも少ない方が好ましい。
もう一つ「バケツ理論」と称せられるモノがある。カール・ポパーが科学論における帰納法を批判して出してきた概念である。簡単に言えば、情報の海の中に穴(感覚器官)の空いたバケツ(ヒト)がある。この穴を通して流れ込んだ情報が知識となるとする帰納法に対し、ポパーは主観としての問題意識を欠いた観察は存在せず、主観による問題意識を通して認識されたものが「開かれた」批判的討議を通して客観的な知識へと成長すると主張した。そして帰納法をバケツ理論と名付けて否定すると同時に、主観としての問題意識をサーチライトに例えた「サーチライト論」を提唱した。こちらのバケツなら、理解可能である。
何で急にバケツかと不審に思われるかもしれないが、実は私も「バケツの穴理論」というモノを考えていた。ポパーが言うところのバケツはヒトである。バケツの穴とは感覚器官である。そして、このバケツが情報の海にある。ここまでは、私の考えとよく似ているが、ここからが違う。私の言うバケツの穴はきわめて小さい。そのため、流入する情報は穴の大きさで制限され、外界にある殆どの情報は認識できないという理論である。
少し具体的に述べるとすれば、我々が聞くことのできる音は個人差があるとはいえ、20Hzから20,000Hz程度である。もはや私には、12,000Hzあたりからのモスキート音は聞こえない。従って、コンビニの前に私が座ったとして、モスキート音による嫌がらせは成立しないだろう。20Hz以下の音および12,000Hz以上の音に対して、私はツンボであると言うことだ。いかに豊穣な世界がそこにあるにしても、私にとっては存在しない。
同じく、我々が認識できる電磁波の範囲は、380 nmから750 nmの範囲である。380 nmより波長の短い電磁波は、紫外線、X線、ガンマ線と分類されており、750 nmより長波長の電磁波は赤外線、サブミリ波、ミリ波、センチ波、極超短波、超短波、短波、中波、長波、極超長波と分類されている。勿論、これより短い波長の電磁波もあれば、長い波長の電磁波もあるだろう。ともあれ、上記の分類も恣意的であり、出典によって異なる分類が存在するとはいえ、γ線の周波数は1020~1023Hz(波長は10-12~10-14 m)、超超長波の周波数は3 Hz以下(波長は108 m)程度であろう。この範囲で我々が認識できるのは0.75 ×10-6 mから0.38×10-6 mというきわめて狭小な範囲に過ぎない。
いま、この空間にはラジオ、テレビ、各種レーダーなどの電波が飛び交っている。電子レンジから漏れたマイクロ波も携帯電話から発信される極超短波も、それらの全てが、我々にとっては存在しない。(本当は、これらの電波は有害であるのかもしれないが) ここにおいて、赤外線健康炬燵のカバーに赤い繊維を貼り付け、ランプから出る赤色光を強調するという感覚詐欺的行為が成立する。されど、我々の視覚に対する信頼は盲信に近い。百聞は一見に如かずという諺通り、見ることが信じることの第一歩となっている。しかしながら、百見しても決して見えない広大な情報の海が存在しているのである。
我々は、バケツにあいた小さな穴から漏れてくる情報のみをもとに、決して見えない外界を考えているに過ぎない。科学の進歩とは、我々が感知できない情報を、いろいろな機器を用いて可聴域の音と可視域の電磁波に変換することによって成し遂げられてきたとも言えるだろう。何が言いたいのか? 我々は、花を我々の目に見える色によって認識し、その色の意義を解釈してきた。この花は赤い、その花は黄色い、あの花は青いと。さらに、その色の組み合わせが、ヒトにとって心地よいモノであれば、これを綺麗、艶やか、華麗、可憐などと評するだけでなく、虫たちの行動にもその評価基準を押しつけ、説明してきたにすぎない。つまり、我々が感知できる可視域での情報のみに従って解釈してきたわけである。この判断態度は「バケツの穴から大海を覗く」が如き見識の狭さに立脚している。では、可視域での情報を越えた判断は可能か?
可視域を越えた立場から虫たちの行動を解明した例としては、小原嘉朗氏らによるモンシロチョウの翅の紫外線反射率による雌雄の識別が良く知られている。花の紫外線写真から昆虫の行動を推測するような研究などは、結論の妥当性は別にしてそうした試みに当たると考えて良いだろう。では、ここで問題にしている花色についてどのような推論が可能か?
フラボノイドに関してはその吸収する紫外線の波長がUV-Bの波長域に相当するため、植物を保護していると考えられているが、カロテノイド類やベタレイン系色素に関しては、それらの持つ可視域の強い吸収帯に幻惑されて、紫外域に関する議論は殆どなされていないようだ。例えばβ-カロテン、この化合物は450 nm付近にεmax (モル吸光係数) = 140,000という極めて強い吸収を持つ。このピークを記録用紙に納めるように測定の感度と濃度を調節すると、280 nm付近の極大吸収は小さなピークにしか見えない。しかしながら、実際のモル吸光係数は52,500という大きな値になる。さらに二つのピークが非常にブロードなピークであるため、300-350 nm付近の最も吸収の低い部分でもそのモル吸光係数は30,000近い値になり、この値はフラボノイドに匹敵する値である。これはβ-カロテンだけの話ではなくζ-カロテン、ニューロスポレン、リコペンなど植物に広範に分布するカロテノイドにおいても同様なことが言える。
後述するが、フラボノイドはカロテノイドに比べ生合成の開始時期は大きく遅れる化合物群で、陸上植物にしか分布しない。オゾン層が成立した後、植物の上陸に際してフラボノイドが紫外線に対する防御物質として働いたとする説に異論はないが、紫外線が透過する水際までの水生植物の進出を可能にしたのはカロテノイドではなかったか。植物の上陸以前から、光合成のメリットと光毒性のデメリットの間で、植物のきわどい生存を支えてきたカロテノイドの存在意義に関しては、いかに強調してもしすぎることはないだろう。そして今ひとつの花色色素ベタレイン類においても、可視域での強い可視吸収帯とともに、320 nm前後にブロードな吸収帯が存在している。従って、この色素群も、多分ではあるが紫外線に対する防御物質としての機能を持つことは容易に推察できる。ただ、植物における分布が狭く生合成開始がかなり遅れる新しい化合物群であるため、植物の上陸時での貢献はないと言えるだろう。
結論を書く。シアノバクテリアを内包した植物は酸素発生型の光合成能を獲得した。従って、光合成を行う細胞において酸素濃度は非常に高くなることは必然である。さらに、酸素の毒性は、光(紫外線を含む)の照射に伴い増大することも事実である。一方、生殖を担う細胞における酸素濃度の増大は、突然変異の可能性を高めるが故に可能な限り避けることが好ましい。従って、花という生殖を担保する器官において、葉緑体を失うという進化は非常に有益であったと考えて良い。そのような進化は、捨てられることなく、現生の植物にまで保持され続けたのであろう。さらに、葉緑体を失った器官において、太陽光照射に伴う紫外線傷害の軽減・除去ができれば、遺伝子変異の可能性をさらに低下させることが可能となる。つまり、葉緑体を失った生殖器官が、障害を起こす紫外線の波長域に吸収を持つカロテノイドやフラボノイドを維持し続けたことが極めて好ましいことは云うまでもあるまい。
ヒトは紫外域での現象をみる能力を欠如していたが故に、UV吸収を持つ化合物群の可視域での振る舞いに幻惑されて、赤だ、黄色だ、青だという議論を続けてきたのであろう。結論は一つ、穴の開いたバケツであるヒトは、穴を通らない紫外線に気付くことなく、穴を通った可視域の光のみを見て、艶やかだだの奇麗だなどという的外れの議論をしていたのである。こうした人の行動に対して「バケツの穴理論」と揶揄したのである。
「毎日、五月晴れの下、山畑での作業をしているのだが、湧き上がる入道雲のような照葉樹の新芽が実にきれいである。濃い緑ではなく、どの木の新芽も緑は薄い。ふと考えた。そういえば、植物の新芽、芽立ちの部分は葉緑体の少ないものが多いと。考えて見れば、植物の新芽、まだ生長を続けている柔らかい部分では、細胞分裂が盛んに行われている。こうした部分において活性酸素の濃度が高ければ、細胞機能に損傷を受けるだけでなく、突原変異の可能性も上がるに違いない。植物が、淡緑色の新芽のだけでなく、赤い新芽(アカメカシワやカエデの仲間など)、黄色い新芽(黄金マサキ)などを持つ理由について、先の結論を適用すれば、その理由がシンプル且つ合理的に説明できるではないか。」 「」で括った部分は5月に書いた。いまの季節には一寸合わない。しかし、こうした問題意識を持って周囲を見渡せば、剪定を追えた後に出てきた新芽も色が薄い。(書いた時期を反映しているので、3月に書いた訳ではない)
以前に植物の色素について述べた。「人間、バケツの穴理論」のことである。我々が感知できる可視域の光にかまけて、紫外域の光を見落としてきたのではないかという内容であった。この議論の中で、煩瑣になることを恐れて素通りした項目がいくつか存在する。それについて少しだけ付け加えておきたい。
紫外線写真というものがある。紫外線フィルターといえば、通常は紫外線をカットして写真の青みを抑え、可視域でのピントをシャープにするために使用する。これに対し、紫外線写真とは、可視光線と赤外線をシャットアウトして、反射してくる紫外光を使って写真を撮ったものである。当然の話として、可視光線で採った写真とは異なった画像がえられる。以下に二つのホームページのアドレスを示す。
1. http://www.fukuoka-edu.ac.jp/~fukuhara/uvir/hana_uv.html:このページには紫外線写真の取り方を含めいくつかの花の紫外線写真が紹介されている。
2. http://www.naturfotograf.com/UV_flowers_list.html :多種類の植物の花の可視光での写真、紫外線写真、蛍光写真、赤外線写真など、豊富な写真が見られる。
紫外線部分を使って花の写真を撮ると、当然ながら可視光で撮ったものと違う模様が現れることがある。この現象は、植物の花の部分におけるUV 吸収を持つ物質あるいはUVを反射する物質の分布を反映するものである。私は先に述べたように、花における葉緑素の消失と、花色色素であるカロテノイド、フラボノイドそしてベタレインの存在を、UV光、可視光を含めた光傷害(直接的な核酸やタンパク質に対する傷害と光照射に伴い発生する活性酸素傷害)からの防御の観点から好ましかったであろうと考えている。もう少し正確に言えば、「葉緑素を持たず、かつ有害な紫外部と可視部で光を吸収するカロテノイド、フラボノイドそしてベタレインを含む花を持つ植物のほうが、有害な突然変異が少なかったであろう」と考えている。
一方、この模様を虫や鳥に蜜のありかを教える印だとして解釈する考え方がある。蜜標(Nectar guide)という概念をもって、虫や鳥が紫外部で見ると黒く見える部分に導かれると説くのだが、余りにも筋の悪い仮説のようだ。花の中心に黒く写る部分があると、それは蜜のありかを示すシグナルであると説明し、アルストロメリアやツツジのように花弁上に黒い部分があると、それは昆虫が着地する位置を示す役割を持つという。そうした吸収パターンを持たない植物も多数存在する。多くの虫を集めるヤブカラシの花には特徴的なパターンは存在しない。花全体がUV吸収を持つ花もあれば、UVを反射する花もある。この辺りの議論については、次の和文の文献を見て欲しい。
福原達人(2008a)植物形態学、紫外線透過フィルタで撮った花。
http://www.fukuoka-edu.ac.jp/~fukuhara/uvir/hana_uv.html
山岡景行、文系学生のための生物学教材の改良、IV:被子植物の蜜標、その2 蜜標の擬似紫外線力ラー画像、東洋大学紀要 自然科学篇 第53号 69-87(2009) http://id.nii.ac.jp/1060/00002546/
山岡氏はその中で、「筆者の自宅がある千葉県柏市郊外ではホシホウジャクMacro-glossum pyrrhostictaが、オオマツヨイグサが開花する時間帯である日没前に花々を求めて庭を乱舞する。同種はこの時間帯にメドウセージSalvia gztaraniticaやチェリーセージS.microphylla、オオマツヨイグサに群がる。日没後を過ぎるとオオマツヨイグサを訪れる昆虫は、ブドウスズメAcosmeryx castaneaやオオマツヨイグサを食草とするベニスズメDeilephila elpenor lewisiiに交替する。黄昏時ならばいざ知らず、日没後で灰かに見えるだけのオオマツヨイグサに刻まれた蜜標紋様が役に立つとは考えにくい。
ミツバチを使ったFrish達の鮮やかな実験の印象が強すぎて、訪花昆虫が全てRを見えずにUVを見ると考えがちであるが、これは問題でありUVだけに着目するのは危険である。」
と書いている。論文中の「Rを見えずに」の部分は意味不明だが、「赤外あるいは可視域での光を見ずに」という意味だと解釈した。そう考えれば、彼の発言に同意できる。UV域での像だけでなく可視域での像も含めて議論すべきであるし、彼が述べているとおり誘引性を持つ、花の香気成分も当然考慮すべきであろう。私に云わせてもらえば、花のサイズも考えるべきである。西洋アブラナ(菜の花)の中心部分に紫外線写真で黒く写る部分があり、これが蜜標だと云われても納得できない。ミツバチと花のサイズはほぼ同じで、ハチが止まればその下に蜜腺がある。止まった状態で蜜標がなければ蜜腺にたどり着けないそんな馬鹿なミツバチなどいるはずがないだろう。
さらに、植物は花以外の部分からでも蜜を分泌する。花外蜜腺と云われているもので、ソメイヨシノやアカメガシワ、カラスノエンドウ以外にも多くの植物に存在する。どう説明するのだろう。そうか、アリ植物という概念があった??
http://www.agr.kyushu-u.ac.jp/lab/ine/ueno/attract2.html
http://www.fukuoka-edu.ac.jp/~fukuhara/keitai/kagaimitsusen.html
さて、世の中には擬似相関(Spurious relationship)と呼ばれる相関のない関係が存在する。2つの事象間に相関がないにもかかわらず、いかにも因果関係がありそうに見える場合を指す。「ガスストーブの使用時間が長くなると脳卒中患者が増える」とか、「朝食を規則正しく食べると成績がよくなる」「髪の毛の長い小学生は言語能力が高い」「アイスクリームの消費量が増えると水死者が増える」など、色々な例がある。
多くのデータがあるとき、恣意的にある傾向を持つ例だけを抽出すれば、どんな結果でも導くことができるということだろう。科学を志すものとしては、決してやってはいけないことである。紫外線写真の解釈においても、肉眼で見えないものを初めて見つけたという喜びと興奮があったことは理解できるが、蜜標(Nectar guide)という概念を提案するについては、もう少し冷静にかつ理性的に考えるべきであったと思う。
山岡氏も、あそこまで蜜標(Nectar guide)という現象に疑問を呈したのであれば、一歩進めてこの概念を否定しても良かったのではないか。紫外線吸収パターンに基づくNectar guide という概念の前に、ハチが食料である蜜と花粉を見つけるためにFlower guides を利用するという先行する概念があったとはいえ、検討した母集団の選択が余りにも不適切であろう。私は過激だから、Nectar guideを含む Flower guidesというアドホックな定義そのものを否定する。
植物が蜜の分泌をはじめたのと、昆虫が蜜の利用をはじめたことを時系列で並べれば、前者が先行することは論を待たない。とすれば、なぜ植物は蜜を分泌するのかという問いにまず答えなければならない。殆どすべての植物が蜜を分泌するとすれば、蜜の分泌をしなければならない必然が植物側にあると考えるべきである。その分泌された蜜に対して、昆虫がどう絡んでくるかは、次の段階の問題である。植物は虫を呼び寄せるために蜜を分泌したのではなく、植物が蜜を分泌したから虫がよってきたのである。
分からないものを分からないままにおいておくことに、たまらない不安感を感じる人がいるらしい。私にはその傾向はない。分からない事柄について記憶はするし考えることを止めはしないが、時には人智を越えたものだとして残しておくことが少しも苦痛ではない。考えるネタがあるということは幸せではないか。アリ植物という概念においても、Nectar guideの場合と同種の時間錯誤が起こっている。(https://bamboolab.yamasatoagr.com/wp/wp-admin/post.php?post=5269&action=edit参照のこと)
近頃、植物が蜜を分泌する理由が分かってきたような気がしている。物理学における統一理論みたいな話になるが、植物における色々な現象を包括的に説明できそうである。この話は、章を改めて近いうちに論じることにする。