
解糖系についての序章
大学で生物系の学科を卒業した人だけではなく、高校で生物学を履修した人であれば、少なくとも解糖系という名前だけは聞いた記憶があるだろう。しかしながらこの解糖系、知名度は抜群であるにもかかわらず、きちんと説明できる人は少ない。この経路は生物の基本的代謝系である、絶対に覚えろと言われたから試験前に覚え、試験が終わったら直ぐに忘却したと言う人が大部分だろう。現実の問題として、我々にとって生活にさほど役立つ知識ではないし、覚えていたからと言って飲み屋で披露できる知識でもない。
初めから脱線するようだが、系と経との使い分けで悩んでいる。代謝系と使う場合は系、代謝経路と使う場合は経を使う。系を英訳するとsystem、経はpathwayとなるのだろうが、系路という使い方もある。さらに、径路という言葉もある。径路と経路は同義であると考えて良いと思うのだが、この径路は生化学分野では使われないようだ。今までの経験からすると、解糖に関する用語においては系が使われる。ところがペントースリン酸経路の場合は系ではなく経を使うようだ。どうもいま一つ分からない。多分、使われ続けてきた歴史が反映しているのであろう。軽々に決められない系路と経路、とりあえず著者の感性で使い分けることにする。
そこで解糖系、実生活で役に立たないからといってどうでも良い系であるかと云えばそうでもない。生化学という学問において、この系はとても、いや最も重要かつ基本的な系として扱われている。日本薬学会という薬学の研究者が構成している学会があるのだが、その薬学会には用語解説のページがある。薬学会自体を批判するつもりはないが、このページには常識的な解説が書いてあるので、少し引用することにする。このサイトにおいては解糖系を以下のように説明している。
解糖系
glycolytic pathway, エムデン-マイヤーホフ経路
グルコースを分解して、ピルビン酸や乳酸を生成する代謝経路。大腸菌からヒトまで多くの生物種に保存されている。解糖系に関与する酵素は、哺乳動物ではすべて細胞質ゾルに存在する。反応全体の収支は、グルコース+2NAD++2ADP+2リン酸→2ピルビン+2NADH+H++2ATP+2H2O、もしくは、グルコース+2ADP+2リン酸→2乳酸+2ATP+2H2O、となる。解糖系は、酸素がまったくない状態でもATPを供給できる特徴をもち、激しい運動時など酸素欠乏時の骨格筋(主として白筋)では必須となるほか、赤血球や神経細胞では唯一のエネルギー供給経路となっている。好気的条件下にある多くの組織では、ピルビン酸からアセチル-CoAが生成し、解糖系はクエン酸回路へ基質を供給する経路としての役割を果たす。解糖系の反応の大部分は可逆的であり、糖新生でも同じ酵素が逆方向の反応を触媒するが、ヘキソキナーゼ(グルコキナーゼ)、ホスホフルクトキナーゼ、ピルビン酸キナーゼの反応は、生理的に不可逆であるため、糖新生では別の酵素が触媒する。グルコース以外にフルクトース、ガラクトース、マンノースやグリセロールも解糖系に回収されて代謝される。また、グルコース6-リン酸はグリコーゲン代謝やペントースリン酸回路などの分肢点となるとともに、ピルビン酸からはアラニンが生合成される。(2005.10.25 掲載)(2009.1.16 改訂)(2014.7.更新)
もう一つ、私がよくお世話になっているバイオインフォマティクス研究用のデータベースから引用しよう。KEGG(Kyoto Encyclopedia of Genes and Genomes:京都遺伝子ゲノム百科事典)と言うサイトで、京都大学の金久實教授達のプロジェクトとして構築され、ウェブ上に公開されているものである。そのグリコリシスのページの説明文を引用する。
Glycolysis is the process of converting glucose into pyruvate and generating small amounts of ATP (energy) and NADH (reducing power). It is a central pathway that produces important precursor metabolites: six-carbon compounds of glucose-6-phosphate and fructose-6-phosphate and three-carbon compounds of glycerone- phosphate, glyceraldehyde-3-phosphate, glycerate-3-phosphate, phosphoenolpyruvate, and pyruvate.
訳文
グリコリシス(解糖系)はサイトゾルに存在し、ブドウ糖をピルビン酸に変換し、少量のATP(エネルギー)とNADH(還元力)を生成するプロセスである。グリコリシスは重要な代謝前駆物質である6炭素化合物:ブドウ糖6リン酸、果糖-6-リン酸、そして3炭素化合物:グリセロンリン酸(1,3-ジヒドロキシアセトンリン酸)、グリセルアルデヒド-3-リン酸、3-ホスホグリセリン酸、ホスホエノールピルビン酸とピルビン酸を生産する中心系路(Central pathway)である。
注:このKEGG(Kyoto Encyclopedia of Genes and Genomes:”京都遺伝子ゲノム百科事典” )というサイトは、1995年に京都大学化学研究所の金久實氏らによるプロジェクトとして発足したあと、現在も整備が続けられている膨大なデータベースである。極めて有用なサイトである。その中のグリコリシスの説明文に異論があるからといって、その存在価値は些かも揺るがないことを付け加えておこう。こんな文章が書けるのもKEGGのお陰である。
上記の説明を読んで即座に「うむ」と納得できる人は極々少数であるとと思う。大多数の人が、科学者と呼ばれる集団が使う専門用語にたじろいでしまうのではないだろうか。もう少しわかりやすくならないかと思わないでもないが、これはこれで仕方ない。専門用語を一応理解した上で記憶し、論理的に使いこなせるようになることは、専門分野を修得するに際して避けて通ることは出来ない。さらに、これらの内容が判りにくいからと言って、その記述が間違っているわけではない。これらの説明文を書いた筆者たちの住むパラダイムの中にあっては、これらの説明は間違いではないのである。
では、先の二つの説明の何処に異論があるのかと問われるのだろうが、一言で答えるのは甚だ難しい。詳しい話は後ろに回すとして、一つだけ疑問をを投げ掛けておこう。確かに、解糖系は大腸菌からヒトまでどころではなく、原核生物(古細菌を含む)から真核生物にわたる多くの生物に広く分布する代謝系である。その点に疑問はないのだが、出発物質がブドウ糖であることに納得が行かない。この系を持つすべての生物は、エネルギー源として使うブドウ糖の給源を何処に求めているのだろうか。さらにだが、解糖系らしき系を持つ何種かの生物群には、その系の中にブドウ糖を含まない生物も散見される。それらをどのように説明すればいいのか?
本格的な批判をする前に、解糖系について常識的な説明しよう。現在認められている解糖系についての知識がなければ、批判が批判ではなくなってしまうからである。さらにだが、この批判は次に述べる「TCAサイクルに関する異論」と続けて読んでもらったほうが理解しやすいだろう。現在、原稿に手を入れているので請うご期待というところである。そこで解糖系、解糖系は図1のように描かれるのが通常である。まあ縦に化合物を並べる場合もあるが、内容は同じである。そこで、この図に沿って説明をする。
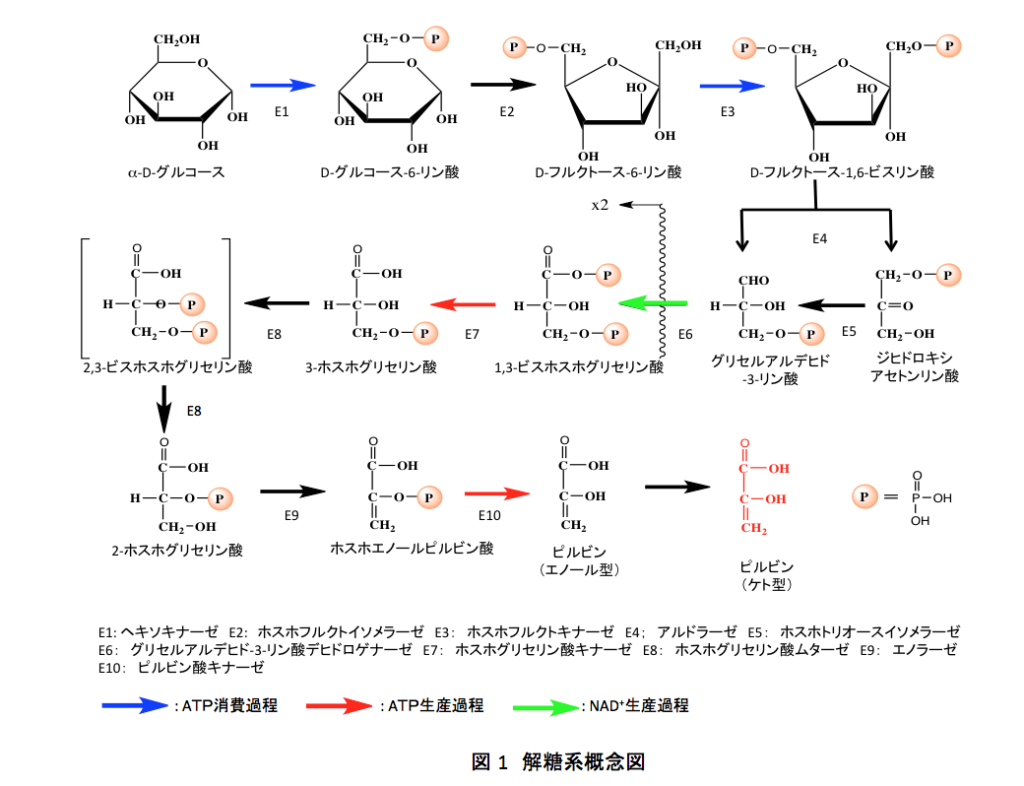
解糖系の出発物質であるα-D-グルコースは、ヘキソキナーゼの触媒下にATPを消費してα-D-グルコース-6-リン酸となる。生成したα-D-グルコース-6-リン酸はホスホフルクトキナーゼによってD-フルクトース-6-リン酸へと異性化される。D-フルクトース-6-リン酸はホスホフルクトキナーゼによってATPを消費しながら、D-フルクトース-1,6-ビスリン酸へと変えられた後、アルドラーゼの触媒下に3位と4位の炭素間の結合が開裂し、3単糖であるグリセルアルデヒド-3-リン酸とジヒドロキシアセトンリン酸が生成する。ジヒドロキシアセトンリン酸はホスホトリオースリン酸イソメラーゼによりグリセルアルデヒド-3-リン酸へと異性化されるため、ここから先は2分子のグリセルアルデヒド-3-リン酸が系を流れることになる。
グリセルアルデヒド-3-リン酸はグリセルアルデヒド-3-リン酸デヒドロゲナーゼによって基質レベルでの酸化を受けた後、リン酸を取り込んで1,3-ビスホスホグリセリン酸となる。混合酸無水物である1,3-ビスホスホグリセリン酸は高エネルギー化合物であるため、ホスホグリセリン酸キナーゼの存在下にADPをリン酸化してATPを生成しながら3-ホスホグリセリン酸に変化する。後で議論することになるが、この3-ホスホグリセリン酸は解糖系を考える上で重要な役割を果たす化合物である。記憶に残しておいて欲しい。
3-ホスホグリセリン酸から2-ホスホグリセリン酸ヘの変換はホスホグリセリン酸ムターゼで触媒されるのだが、この酵素には2種類が存在する。一つは植物、古細菌そしてグラム陽性菌を中心に分布する酵素で、分子内でリン酸基の転移を触媒する酵素である。いま一つは、脊椎動物、昆虫、藻類、そしてグラム陰性菌に主に分布する酵素で、2,3-ビスホスホグリセリン酸をコファクターとして3-ホスホグリセリン酸の2位の水酸基のリン酸化に続く3位のリン酸残基の脱離を通して2-ホスホグリセリン酸を生成する。図1には、後者のタイプの反応をイメージして、2,3-ビスホスホグリセリン酸を中間体とする形で示している。
こうして生成した2-ホスホグリセリン酸はエノラーゼによって脱水反応を起こし、極めて重要な代謝中間体であるホスホエノールピルビン酸に変換される。ホスホエノールピルビン酸はエノール型になったピルビン酸の水酸基がリン酸化された化合物で、ピルビン酸キナーゼの存在下にADPをリン酸化してATPを生産しながらエノール型のピルビン酸にとなるのだが、このエノール型のピルビン酸は極めて不安定で触媒の存在を必要とせず速やかにケト形のピルビン酸へと異性化する。
解糖系においては、青の矢印で示した2つの段階でATPが消費され、赤の矢印で示した2つの段階でATPが生産される。グリセルアルデヒド-3-リン酸以降は系を2分子が流れることより、解糖系を通ってα-D-グルコースが2分子のピルビン酸に分解されると差し引き2分子のATPが生産されることとなる。また、緑の矢印で示した段階で、NAD+が還元を受け重要な補酵素NADH2分子が生成する。
つまり、「ほとんどの生物が持つ嫌気的でもっとも普遍的かつ根源的な代謝系」である解糖系は、6炭糖であるブドウ糖を出発物質とし、酸素を使わずに2分子のピルビン酸まで分解することにより、2分子のATPと2分子のNADH2を生成する反応である」となる。
学生の立場からすれば、「解糖系について知見を述べよ」という問題は、図1を記憶して、解糖系が「ほとんどの生物が持つ嫌気的でもっとも普遍的かつ根源的な代謝系」であること、2分子のATPと2分子のNADH2を生成する反応であることを書けば、間違いなく80点は貰える楽勝な問題である。系を乳酸まで伸ばせば、NADHの収支もゼロとなり、細胞内の酸化状態も変えずにATPの生産ができると書けば、今少しの加点が期待できる。私だって学生の時はそう答えた。現役教員の頃もそういう基準で評価をしていた。失礼ではあるが、分かっていないのにこの図を丸暗記する集中力と努力は認めるべきだと考えたからである。こいつ何にも分かっていないくせにと思っていたにせよである。
本論から逸脱することは承知の上で、少しだけ補足しておきたい。代謝系を構成している反応群は、少なくとも有機化学的に理解されるべきだという立場にいる私から見ると、図1をとにかく丸暗記するという時点で理解することを放棄したと言える。これは当時の学生達を揶揄しているのではなく、学生時代の私自身に対する批判でもある。
私も図1のような概念図を絵として暗記し、それを答案に書いた。単位は取れたのだが、何も分かっていないことには気付いていた。有機化学に片足を乗せて研究生活を始めようとしていた学生であるにもかかわらず、この系を構成する素反応群を有機化学的に記述できなかったのである。D-フルクトース-1,6-ビスリン酸とグリセルアルデヒド-3-リン酸・ジヒドロキシアセトンリン酸の間で起こる可逆反応なんて、とても刃が立たなかったし、グリセルアルデヒド-3-リン酸から1,3-ジホスホグリセリン酸への変換はさらに理解不能な反応であった。
つまり、当時の私にとって図1は単なる絵に過ぎなかった。生物有機化学という分野自体がまだ一般的ではなかった時代、多くの先生方さえもが有機化学と生化学は別の学問であるという考えであったようだ。代謝系の素反応群を何とか有機化学的に理解したいと、ドナルド・J・クラムが有機電子論的立場から書いた有機化学の教科書を、練習問題まで全て解きながら5回ほど読み通した。進歩は遅く、解糖系の素反応を有機電子論的に描けるようになるのに5年ほどかかったのだが、そうなってみると新しい世界が広がっていたのである。この頃、1年後輩ではあったが、適切な指摘と助言をくれたK君がいなかったとすれば、今の私は無いだろう。できる後輩を持つというのは半分有り難く、半分怖いものである。感謝!!