
依然として気になっているのはCHS/STSが形成している大きなクラスターの外側にいる2つの配列F6GV80とF6I151である。シロイヌナズナの場合と同様に、偽遺伝子化して進化速度が上がったもう一つの例であり、この配列を除外して解析したのは合理的判断であるとして話を閉じるのが賢いことは分かっている。そうしておけば、植物体内にはCHSあるいはSTSとしての機能を維持しながら、着実に進化を続けてきた配列群と、途中で偽遺伝子化して高い速度で進化してきた配列群が存在すると結論づけることができるだろう。
だがその結論にそこはかとない違和感を感じるのである。私の感覚に従えば、アラビドプシスについての解析結果とブドウについての解析結果が余りにも似すぎている。ゼニゴケのスチルベンシンターゼQ516Y1と一応正常と判断している上部の配列群が4.7億年前に分岐したとすれば、クラスターの外側にいる2つの配列F6GV80とF6I151との分岐時期は10億近い値となる。この10億年程度という値がアラビドプシスの場合とほぼ同じになるのが違和感の原因である。ブドウの異常配列とアラビドプシスの異常配列には何か関連があるのだろうか。
そこで、両者を一緒にして解析て得られた樹形図を下に示したのだが、なんとブドウのはみだしものであったF6GV80とF6I151はシロイヌナズナの異常なCHSと類縁関係にあったのである。いわゆる正常と思われる(現在機能している)グループとは別に、十億年ほど前に分岐した(?)と思われる別グループ−偽遺伝子化に伴う進化速度の昂進で発生した偶発的な配列と考えていたもの−が意外な広がりを持っている可能性が出てきたのである。
若い頃だけではなく研究者としての晩年になってからも、論文を書くのが嫌いだった。研究とは最終目的は動かさないにしても、1段階ごとに得られる結果を基に、次のステップの実験予定を立てるものである。ところが、学術論文と呼ばれるものにおいては、Materials and Methodsという段落において、実験方法と実験材料を先立って書かなければならない。その後 Results and Discussion において結果と考察を書くのだが、どうしても実験の流れと考察の流れにギャップができるのである。ある実験をした。こんな結果が得られた。その結果を基礎として次の実験を行うとこんな結果が得られたと書きたいのである。つまり、“そこで”という接続詞を用いたいのだが、科学論文においてはそうした書き方は認められない。私にとって論文を書くという行為は、自粛、我慢、自己規制と同じだった。正直に言えば書きたくなかった。未熟だった頃一度だけ、こう考えて実験を行ったらこうなった、そこでこういうことが原因だと考え、それを確かめるために次の実験を行った。と言う書き方で投稿したことがある。もちろん没、門前払いであった。その後、何人かの知人たちが論文の書き方をさり気なくレクチャーしてくれた。あいつに論文の書き方を教えてやれという編集委員からの暖かいサジェストがあったと聞いている。気にかけていただき有り難かったのだが、それでも少しだけ反抗したかったのです。
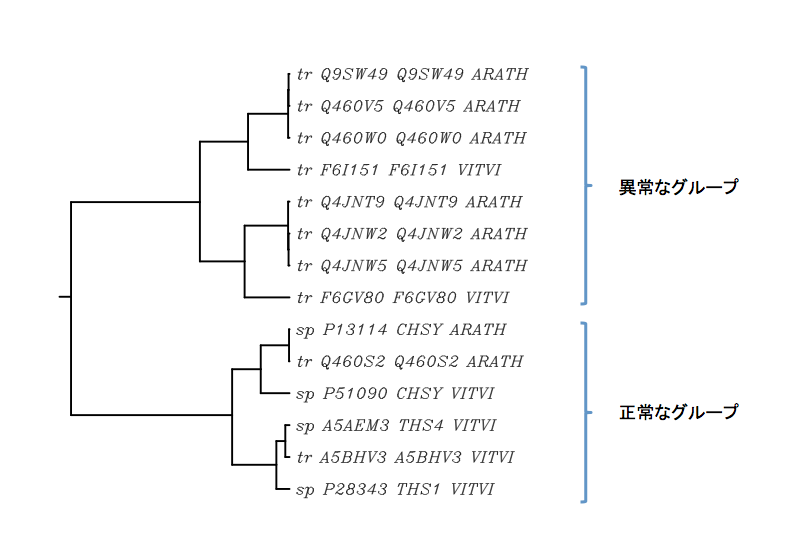
さて、十億年ほど前に分岐した(?)と思われる異常グループ−偽遺伝子化に伴う進化速度の昂進で発生した偶発的な配列群と考えていたもの−が意外な広がりを持っている可能性があると書いたのだが、それはブドウとシロイヌナズナについてだけの暫定的結論に過ぎない。この別グループの配列群は、植物の中でどの程度の広がりを持っているのだろうか。そこで(この「そこで」が自然に使える喜びはなにものにも変えがたい)、CHSの進化速度の解析を行った時に削除した異常と思える配列群を含めて、少しまとめて解析してみたい。定性的な表現で科学論文としては成立しないかもしれないが、各植物の中で異常な配列をもつとして解析から除いた配列の中には、当然非常に大きく異なっているものと、さほどでもないものが存在した。こうした配列群を選び、先に解析したシロイヌナズナの6本、ブドウの7本、さらにゼニゴケのSTSを加えて描いた樹形図を下に示している。

この図において、タンパク質として発現している正常な配列をもつと考えられるQ5I6Y1、P13114およびP28343が属するクラスターに布置される配列群と、これらと全く異なった配列群(ゼニゴケとの分岐より遙かに遠い時期に分岐したグループ)は二つのクラスターにきれいに分かれるのである。そしてこの異常クラスターにはシロイヌナズナとブドウの異常配列群だけでなく、ハクサンハタザオ(Q460X0、Q460X6)、モントレーマツ(O24484)、イネ(Q43595)、パンコムギ(C3RTM5)のもつ配列群が含まれ、植物の中で予想を超える広がりを持つことが明らかになった。
結論というほどのものではないが、私が行った解析が有効なのは、どうやら前の正常としたクラスター内に限られるようである。この正常と思われる配列群からなるクラスターは、植物内でCHS/STSとして機能する配列として進化してきた。もちろん、このクラスターの中にも幾分異常と思われる配列群が存在するが、それらについては偽遺伝子化して急速に進化し、復帰変異によって再度酵素活性を獲得したと考えてよいだろう。
一方、10億年ほど前に分岐した(?)今ひとつ異常のクラスターは、いかなる意味を持つのか? いや、本当に10億年ほど前に分岐したかどうかも分からない。これらの配列群が転写も翻訳もされない、単なる類似シーケンスを持つだけならまだ疑問は小さいのだが、これらの中には転写段階で確認されている配列もかなりな数存在する。さて、それらは植物の体内でどのような意義を持っているのだろうか。10億年ほど前と言えば、真核生物が多細胞化した時代に近い。だからといって、何か説明できるかと言えば何も出来ない。この部分はしばらくの間、謎として残し、時々思い出しては考え続けていくしかないだろう。知的忍耐力が試される訳である。異常が解析において使わなかったデータについての論証です。これを隠蔽と見るかどうかは読者の判断にお任せするしかないだろう。都合が良いか悪いかは別にして、現時点で隠蔽しているデータはなくなった。次へ行こう。