
- 2020.08.16 Sunday
- 01:12
さて、以前に次のように書いた。【そうした立場(β-カロテンの生合成に酸素は必要か?)から眺めるとすれば、poly-cis-carotenoid pathwayとかall-trans-carotenoid pathwayとかいう経路の中で、cisであるtransであるというような小さな差異で議論するのは適当ではなく、プラストキノンをはじめとするキノン類を酸化剤として用いる酵素群[EC 1.3.5.5 and 1.3.5.6]と酸素を酸化剤として用いる酵素群[EC 1.3.99.26, 1.3.99.28, 1.3.99.29, 1.3.99.30, 1.3.99.31]に分けて捉えればよいようだ。】 つまり、phytoene desaturaseを始めとして、連続した脱水素により共役二重結合を生成していく酵素群が、酸化型のプラストキノンを水素受容体として利用して酸素分子の関与なくβ-カロテンの生合成が可能だと述べたわけである。
しかし、よく考えて見るとそれだけでは不十分である。水素を受容した還元型プラストキノン即ちプラストキノールの再酸化はどのように起こるのかを抑えておく必要があるだろう。この問題を詳らかにしておかないとβ-Caroteneが還元的に生合成されるという言明は成立しないことになる。
そこで考えるべきは、現生の高等植物についての話ではない。シアノバクテリアの出現までに成立していたβ-カロテンの生合成系についての話である。勿論、表8-1-1~3に示したように、光合成細菌にβ-カロテンを生合成する種が存在することを傍証とすればよい。ではβ-カロテンを生合成するこれらの光合成細菌は、プラストキノールの再酸化をどのように行っているのだろうか。
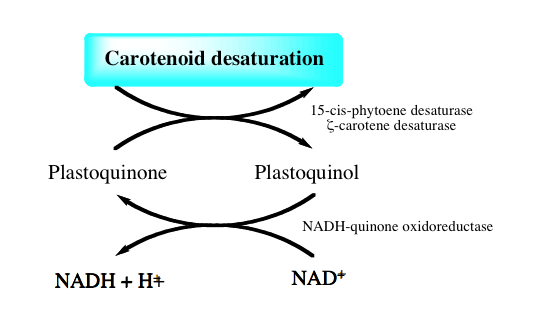
図 8-10 カロテノイドの不飽和化によって生じたプラストキノールは
どのように再生されるか?
調べてみると、図8-10に示すようにカロテノイドの不飽和化で生成したプラストキノールやユビキノンは、細胞中の酸化還元反応で広く使われるNAD+(酸化型ニコチンアミドアデニンジヌクレオチド)をNADH+H+(還元型ニコチンアミドアデニンジヌクレオチド)に変換されるようだ。こうなれば、多くの細胞中の酸化還元反応に共役できることになり、プラストキノンの再生に酸素を必要とすることはない。phytoene desaturaseやzeta-carotene desaturaseによるプラストキノールやユビキノールの生産は、大きなNADH+H+プールの小さな波紋として受容されていくのであろう。この問題は片づいたと考えて良い。
次に問題になるのは、β-caroteneからabscisic acidまでの酸化反応の連続に関する部分である。植物細胞においてアブシジン酸の生合成は色素体つまり葉緑体で起こっている。葉緑体においては非メバロン酸経路が動いていることを考慮すれば、恣意的ではあるがアブシジン酸生合成の出発物質はピルビン酸とグリセルアルデヒド-3-リン酸と考えても大きな異議はでないだろう。そうすると、この2つの物質からアブシジン酸までに約26段階の反応が存在する。なぜ約という接頭語がつくかといえば、段階数の違うバイパスの存在、或いはIPPとDMAPPの異性化をどう数えるかなど厳密に数えにくい段階が存在するからである。
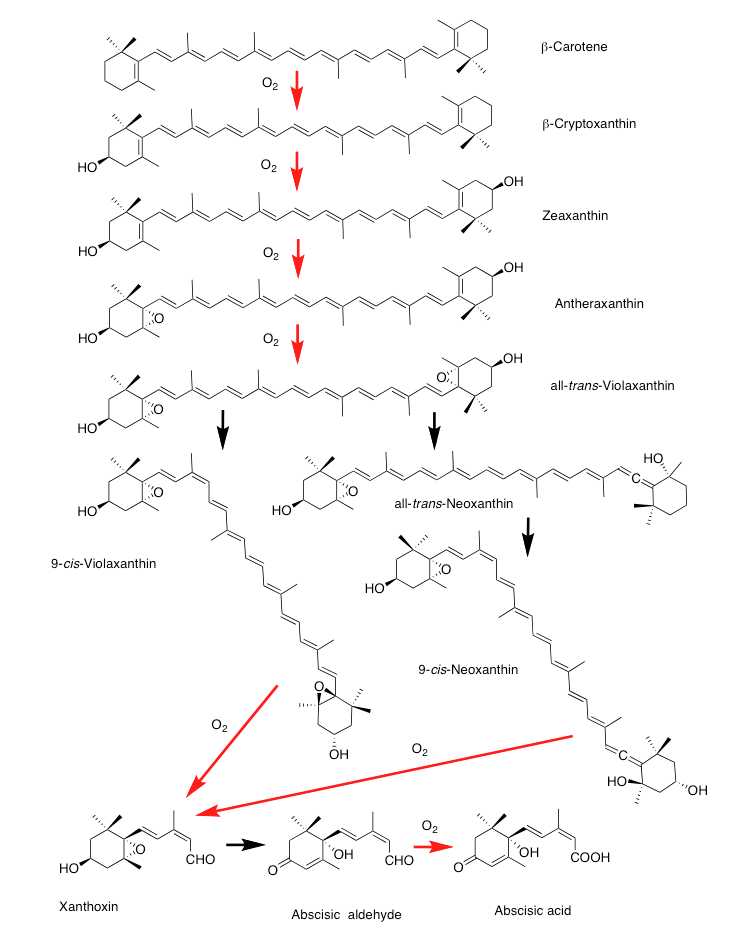
図8-11 アブシジン酸の生合成系で酸素分子が関与する酸化反応
先にも述べたが、この26段階の反応中に酸素が関与する反応は6段階でみられるが、これらは全て、βカロテン以降の反応段階に偏在している。図8-11に示すように、βカロテンからアブシジン酸までの8段階の反応においてall-trans-Violaxanthinから9-cis-Violaxanthinへの酵素さえ分かっていない異性化反応とXanthoxinからabscisic aldehydeへの反応以外、全て段階が酸化反応である。反応の性質上、βカロテンからβ-cryptoxanthin、β-cryptoxanthinからZeaxanthinへの2つの酸素添加反応反応とZeaxanthinからAntheraxanthin 、Antheraxanthin からViolaxanthinへの2つのエポキシ化反応は、それぞれ時を違えず出現したと考えるとしても、4段階の酸化反応の伸長にはどれくらいの時間がかかるのだろうか。
アブシジン酸の分布を思い出してみると、基本的には蘚類以上ということになっているが、緑藻類、シアノバクテリアにおいても微量の存在が明らかにされている。コケ類と緑藻類では、生育抑制的に働くようだが、シアノバクテリアには目立った活性はないようだ。(スピルリナに与えてみたことがあるが、殆ど影響はなかった)植物が上陸したのは約5億年前、シダ植物と高等植物に向かう枝が分岐したのが4億年ほど前である。コケ類については、シダ類からの退行進化によって生じた可能性が指摘されているとはいえ、化石による証拠からみても分子進化学的データからみてもシダ植物より原始的である。コケ植物から高等植物へと伸びる系統樹において、苔類までは生長調節物質としてルヌラリン酸が機能し、蘚類からはアブシジン酸が機能する。そうすると、アブシジン酸が生理活性物質としてニッチを獲得したのは5億年から4億年前の間であると推定できる。
7章で述べたように、カロテノイドの歴史はステロイドより長いと推定出来る。Brocks らの報告によれば、27億年前にステロイドの生合成が始まっていた。β-カロテンまでの生合成はスクワレンまで生合成に先行するとはいえ、分子状酸素を用いた酸素添加反応の開始段階で、ある程度揃うのではないだろうか。こう考えると、β-カロテンの酸化は27億年頃に始まり、5億年頃までにはアブシジン酸に到達していたことになる。この数字の妥当性については、もう少し後で議論することにしよう。
いま一つ、私が感じていた疑問は、「なぜ植物は、C15のアブシジン酸を生合成するのにC40のカロテノイドを経由し、これを分解してC25部分を捨て残ったC15部分の片方を使うというまどろっこしい経路を使うのか。」という疑問である。植物病原菌であるCercosporaやBotrytis の仲間は、C15のファルネシルピロリン酸から直接アブシジン酸をつくると報告されている。しかし、植物は実際にカロテノイド経由でつくる。なぜそんな迂回経路を使うのかという疑問は、学会では成立しにくい問いである。近頃、朧気に答えが見えてきたような気がしているが、誰かクリアーな答えをお持ちの方はいないのだろうか。
さて、ルヌラリン酸は下等植物に、アブシジン酸は高等植物に分布する。多くの人がこの記述で納得するらしい。しかし、私は高等とはなんぞや、下等とはなんぞや、との疑問を捨てきれていない。生物を高等、下等という形容詞でくくることは不可能であろう。どちらも40億年程前に発生した原初の生物の子孫であり、地球上で同じ年月を生き続けてきた生き物ではないかと思ってしまう。
とはいえ、古い時代の形質をより多く残しているのは、いわゆる下等植物であることは間違いないであろう。彼らは、環境変化の少ない状況で大きく変わることなく生きのびてきた生物である。この条件に、生合成における酸素の関与を加味して考えれば、古い生長調節物質がルヌラリン酸であり、新しい生長調節物質がアブシジン酸であると推論してよいだろう。今後の議論においては、古い生長調節物質がルヌラリン酸であり、新しい生長調節物質がアブシジン酸であるとして話を進めることにする。