
歴史的な視座から眺めると色々な疑問が残っている。TCA 回路は、いつ地球上に現れたのだろう?初めて出現したTCA回路は、現在の系路と同じものであっただろうか?それとも、不完全な系であったのだろうか? TCA 回路が好気条件下で働く系路であるならば、嫌気的条件下で生きる生物にはTCA回路は存在するのだろうか、それとも存在しないのであろうか?前回の終わりにそう書いた。
こうした形の疑問を持つ学生は極く少数ではあるが存在する。私もそうだった。彼らは完全な形での答えを求めているのではなく、ただ素朴に疑問を出しているに過ぎない。教える側が、そこは私にも分からないと素直に答えれば、ああ先生にもわからないことがあるんだと悪気の欠片もなく喜び、その先生を軽蔑するのではなく尊敬し始める場合があるのである。そして、これを契機に学問を始める学生もいる。
ここで先生方が面子と沽券に囚われると学生との信頼関係が失われ、取り返しのつかないことになる。今になってわかるのだが、先生という人々が教科書を使って教えることなど大した事ではない。社会に出てさほど役立つ物ではないし、新しいといえる内容でもない。役立つとすれば、前に進もうとする師の後ろ姿くらいかな。学生(弟子)には、師を視界の先に在るものを理解しそれを超えようと汗する態度が求められる。この際、師はそうしたことができる弟子を嫉んではならない。
さて本論に戻るとして、以下に示すこれらの図は、いつものようにKEGGのサイトからの引用である。図3-16は光合成細菌であるChlorobium chlorochromatiiの持つ 回路、図3-17は嫌気性古細菌であるThermoproteus uzoniensisの持つ 回路、図3−18は好気性古細菌であるSulfolobus tokodaiiの持つ回路、そして図3−19がHomo sapiens (Human) 持つ TCA回路と呼ばれる系路である。系を動かす酵素にいくつかの差異はあるにしても、系を構成する化合物は同じであり、註釈なしにこれらの図を見せられて、ヒトの持つ系路とヒト以外の生物が持つ系路が同じ存在意義を持つものではないと主張する人がいるとしたら、その人はかなりな変人だと認定されても仕方ない。私も、長い間同じように見えていた。
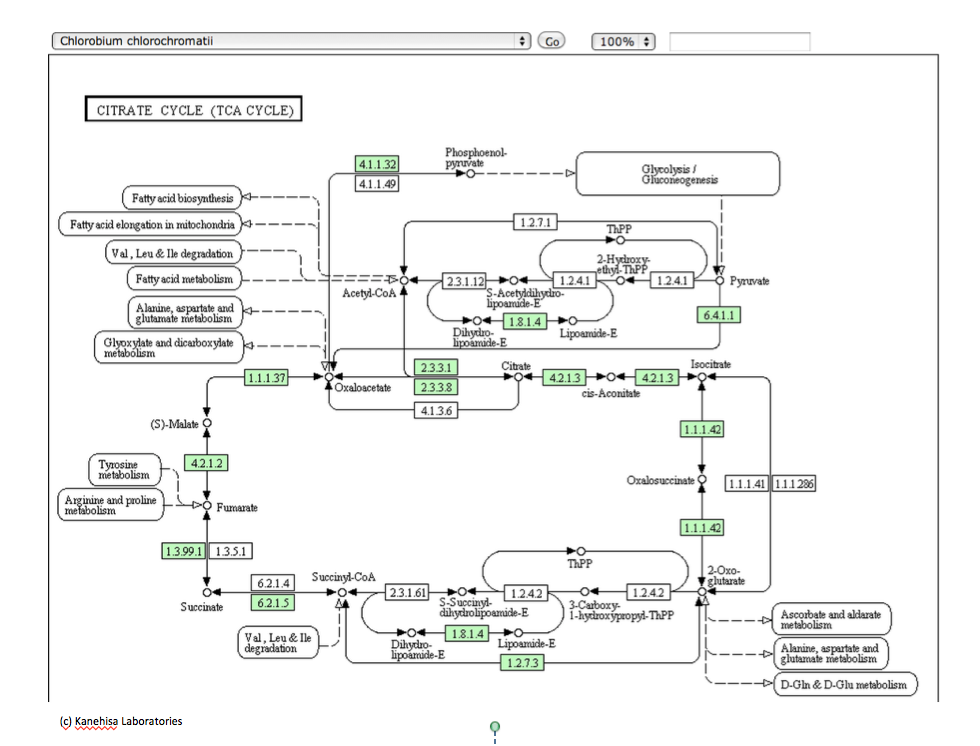
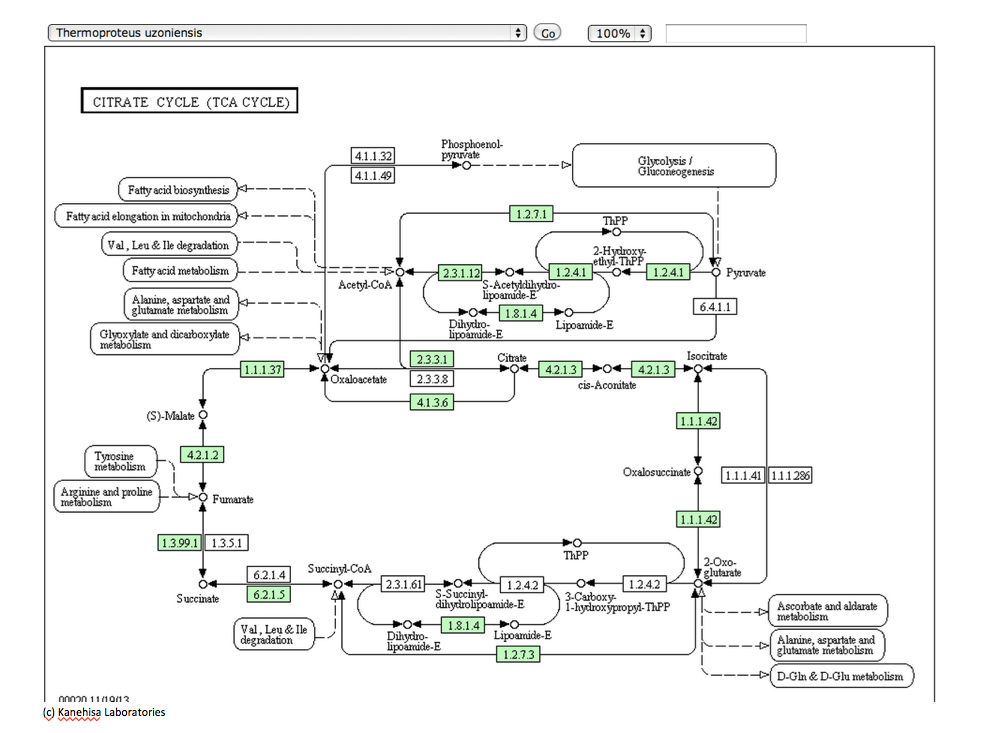
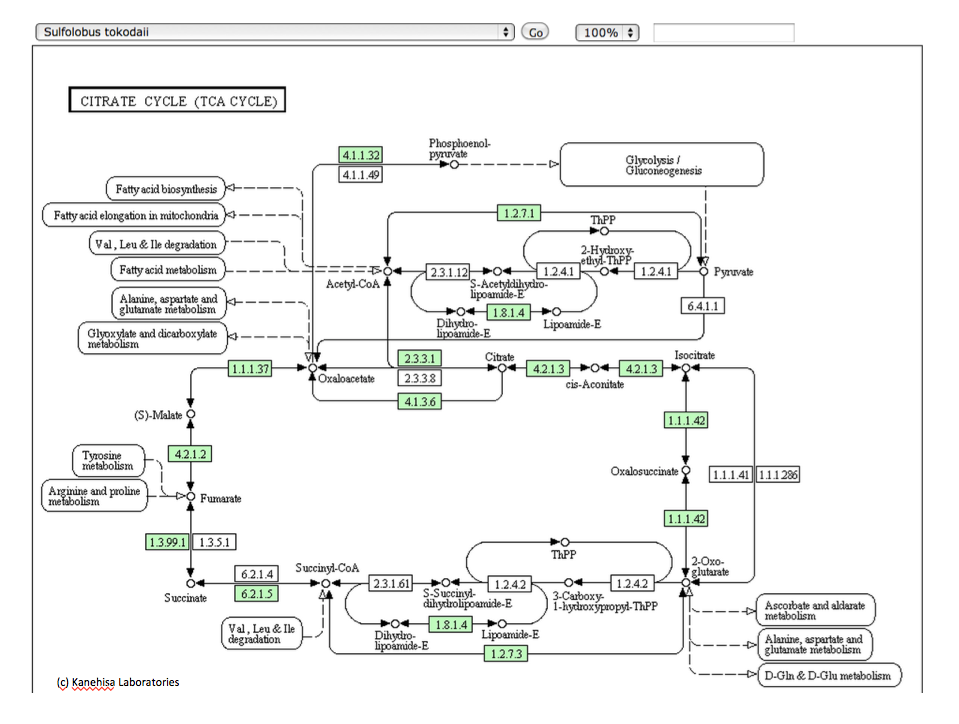
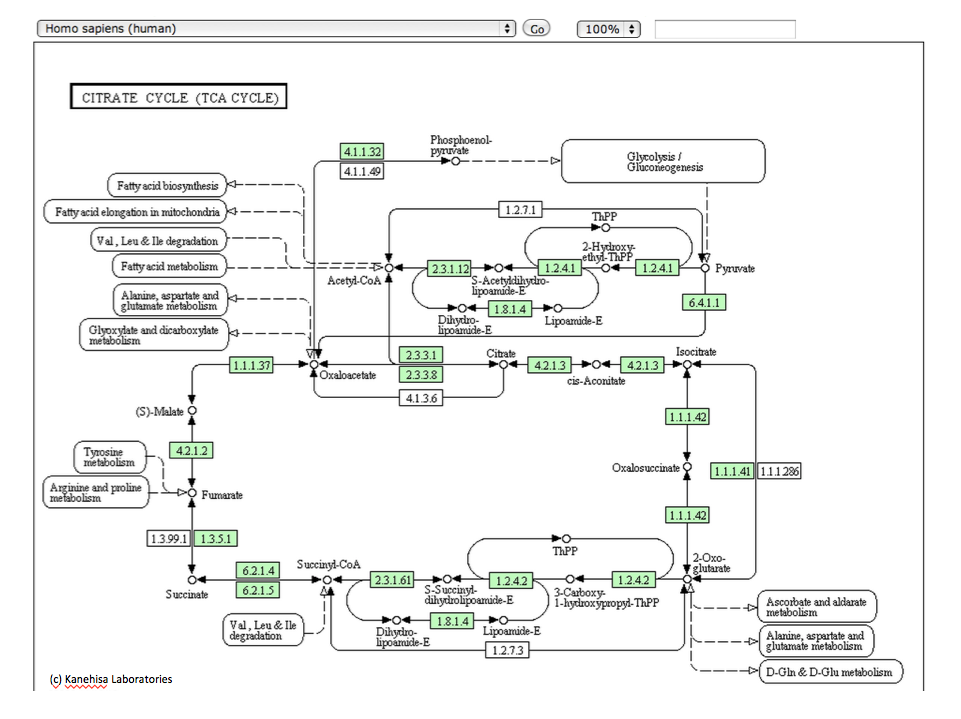
とはいうものの、答えるのが甚だ難しい困った事実が存在する。ヒトを除く上記の3種の生物において、その TCA 回路は右回りに機能しているのではなく左回りに機能しているのである。では、その左回りの系は何をしているのか。図3−2を参照すればすぐに分かることだが、TCA 回路が左回りに機能するとすれば、スクシニルCoA から2-Oxoglutarate、2-Oxoglutarateからオギザロコハク酸の段階で炭素固定が起こっているのである。ある程度以上の知識を持つ人々はこの系を知っており、この逆周りの系を還元的クエン酸サイクルと呼んでいる。解糖系という糖の分解系に、糖新生系という逆向きのベクトルを持つ代謝系が存在したように、TCA 回路にも還元的クエン酸サイクルという逆向きのベクトルを持つ代謝系が併存しているのである。ではこの還元的クエン酸サイクルという逆向きのベクトルを持つ代謝系は何をしているのか。
日本光合成学会が2015年4月に公開したWeb版の光合成事典というサイトがある。その中から還元的クエン酸サイクルという逆向きのベクトルを持つ代謝系についての説明を引用したい。以下 引用である。(https://photosyn.jp/pwiki/index.php?還元的TCA回路)
「 還元的カルボン酸回路,逆転クレブス回路とも呼ばれる炭素同化回路. Evansら(1966)により緑色硫黄細菌Chlorobium limicolaにおいて光合成CO2同化系として存在が提唱され,その後一部の紅色細菌,好熱性水素細菌Hydrogenobacter,嫌気性古細菌Thermoprotenusなどでも存在が報告されている.その起源は還元的ペントースリン酸回路より古いと考えられている.解糖系の一部とTCA回路を組み合わせたような回路で,還元型フェレドキシンを用いるピルビン酸シンターゼによりTCA回路に向かう反応とは逆にアセチルCoAからのピルビン酸合成を行う.緑色硫黄細菌ではPEPカルボキシラーゼによりホスホエノールピルビン酸(PEP)にCO2を固定しオキサロ酢酸を生成する経路が働いている.緑色硫黄細菌ではピルビン酸からホスホエノールピルビン酸への転換をピルビン酸・リン酸ジキナーゼが触媒する.オキサロ酢酸以降はTCA回路の逆転によってスクシニルCoAを生じ, TCA回路では非可逆的な反応であるスクシニルCoAから2-オキソグルタル酸(α-ケトグルタル酸)の合成を還元剤として還元型フェレドキシンを用いる2-オキソグルタル酸シンターゼにより行う.クエン酸の開裂にはATP依存型のクエン酸リアーゼを用いてオキサロ酢酸およびアセチルCoAを生成する.」
TCA回路の説明においては、酸化的リン酸化を含めてエネルギー収支の問題が大きく取り上げられるのに、ここにはエネルギー収支の話は全く見当たらない。何となく不満である。最初にさらっと触れられたように、還元的カルボン酸回路の意義は炭素固定ということで納得して良いのだろうか。
さらにこの逆向きのベクトルを持つ代謝系を持つ生物種の構成を考えると、現在の動物細胞もその細胞質に還元的クエン酸サイクルを持っていたように思える。それが酸化的リン酸化を伴うTCA回路を持ちミトコンドリアの祖先であった好気的細菌との共生の後、還元的クエン酸サイクルを失っていったと考えて良いだろう。つまり好気的なTCA回路が還元的クエン酸サイクルに置き換わった事を意味する。では、好気的なTCA回路は還元的クエン酸サイクルのどの機能を代替したのだろう。還元的クエン酸サイクルがエネルギー生産を行わない以上、エネルギー生産をその意義として置くことはできない、同時に好気的なTCA回路のあるかどうかわからない程度の炭素固定能力にその因を求めるのは、論理的に通用しないようだ。