
要するに、大したことを云っているわけではない。TCA回路と電子伝達系は分けて考えるべきであり、その場合のTCA回路は嫌気的回路と考えないと、解糖系の定義のやり方と矛盾するではないかという指摘をしているだけである。ただし、これを認めると今まで続いてきたTCA回路もしくは解糖系について、どちらかの定義の修正が必要となる。過去に出版されてきた多くの書籍や報告の修正は、JRの駅名の変更のようにとてつもなく大変な作業を伴うのでそんなことは出来ないに違いない。万一認められたとしても、「まあそういう理解もあるよね」というなんとも生温い判断の下で、そっと放置される蓋然性が高い。現段階において私の意見はかなり高い確率で没になる少数意見であることは自認している。とはいえ、こうした異なる視座からTCAサイクルを見たらどうなるのか、もう少し議論を進めてみることにする。
いまひとつの問題は、細胞内でのTCA回路の存在意義に関する定義の部分にある。前章において、人は自らが従属栄養生物であるという実感(一定の時間ごとに空腹感に襲われること)を持つが故に、EM経路という経路をグルコース或いはグリコーゲン(デンプン)を出発物質、ピルビン酸を生成物として流れるATP生産系として捉える考え方に親和性が高いのだろうと述べた。しかしながら、独立栄養生物には流れのベクトルが異なるEM経路が存在する。例えば、植物においてのEM経路は 3-ホスホグリセリン酸が流入点であり、ここから多糖類の生合成へ向かうEM系路とピルビン酸に向かうEM系路が始まっていると考えるべきであると「解糖系への異論」の中で述べた。同じように系を解釈するに当たって、依拠するパラダイムの違いが、TCA回路の意義付けに大きく影響する可能性が存在するのではないだろうか。
TCA回路の存在意義はどこにあるのかという問題に対して、日本薬学会は「好気的な条件下でエネルギー獲得に中心的な役割を果たす」という一文で片付けている。最後の部分で「また、エネルギー的にATPと等価のGTPを1分子産生する」という説明を付しているところを見ると、エネルギー獲得という観点からの判断しか示していない。
一方、KGEEにおいては「In a series of reactions two carbons in citrate are oxidized to CO2 and the reaction pathway supplies NADH for use in the oxidative phosphorylation and other metabolic processes. The pathway also supplies important precursor metabolites including 2-oxoglutarate. (アセチルCoA に由来する)クエン酸中の2つの炭素原子は、一連の反応の中で二酸化炭素へと酸化され、酸化的リン酸化、そしてその他の代謝過程で使用されるNADHを供給する。TCA回路はまた、2-オクソグルタル酸を含む重要な代謝物を供給する。」とのべており、この回路から派生していく他の代謝-特にアミノ酸代謝を意識したような記述となっている。但し、前半でこの回路を好気的過程であるとした記述があるために、TCA回路が好気的回路であるという枠組みは生きているようだ。その点に問題ありと思うものの、KEGGの定義の方に幾分かの柔軟性が認められる。
以下、書くべきか書かざるべきか少々迷った。本筋から幾分離れた内容になるからである。とはいえ、TCA回路を使って動物が二酸化炭素を固定すると書いてあるからには少々コメントせざるを得ない。二酸化炭素を固定するということは、還元するということと同義であり、TCA回路を還元系として捉えることを意味するからである。総合的に見れば、流れる物質量としては少量であり、炭素循環においては無視できると考えて良いかもしれないが、少々気になったので書くことにした。
いまから18年ほど前になる。カイコやクモの糸を構成するアミノ酸中に、空気中の二酸化炭素が取り込まれるという報告が出た。質量数13の炭素を含む二酸化炭素を用いたトレーサー実験の結果であった。13Cを取り込んだ主なアミノ酸はアラニン、アスパラギン酸、グリシン、セリンであるとされていた。13CNMRの積分値を取り込みの証明に使った実験である。13CNMRは、定量性については幾分不確実な(特に完全デカップリング条件での測定)測定法である。感度は低くなるが定量性を重視した測定法もあるためコメントしにくいが、そういうことは織り込み済みでの結果であり結論であると思う。この結論によれば、動物である昆虫が、空気中の二酸化炭素を固定するという結果になり、真核生物である動物は炭素固定を行わないと思っていた私にとって、結構興味を引く報告であった。
ただ、いくらかの疑問は残った。図-9に農業生物資源研究所のサイトに乗っている図を示す。
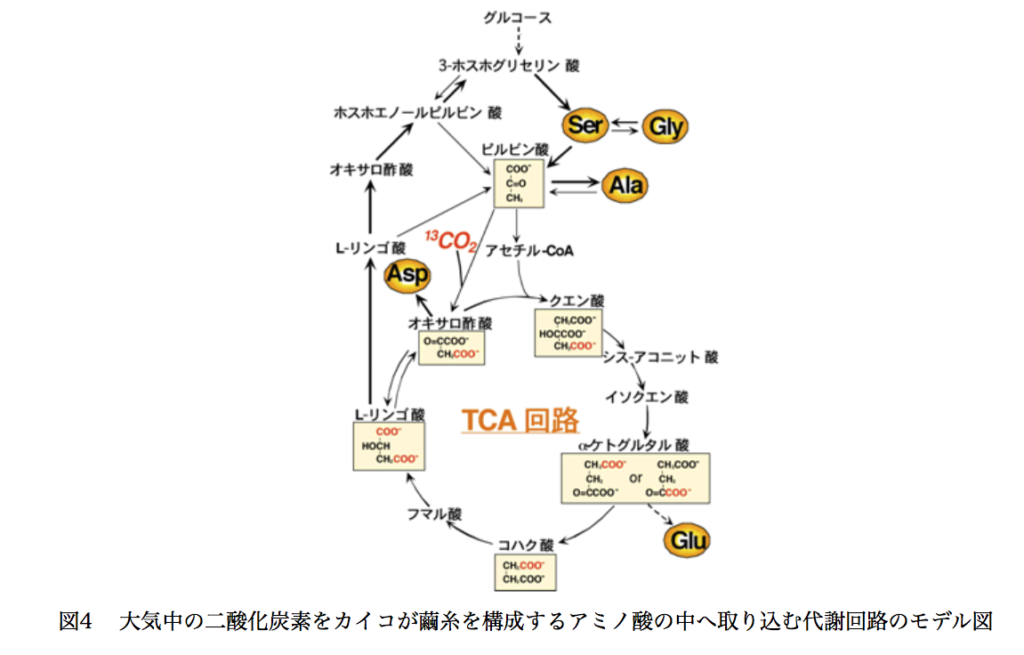
この図に於いては、ピルビン酸が pyrvate carboxylase [EC 6.4.1.1]の触媒下に二酸化炭素を取り込んでオギザロ酢酸になるのだが、この段階で炭素固定が起こっていることになる。オギザロ酢酸は通常通りアセチルCoAと反応してクエン酸を与える。クエン酸から脱水反応が起こってシスアコニット酸に、シスアコニット酸に反対向きの水付加が起こってイソクエン酸となった後、α-ケトグルタル酸に変換されるのだが、カイコには二通りの経路が存在する。一つはisocitrate dehydrogenase (EC 1.1.1.41)によって一気にα-ケトグルタル酸になる経路、いま一つはisocitrate dehydrogenase [EC:1.1.1.42]の触媒下にオギザロコハク酸を経由してα-ケトグルタル酸になる経路の二つである。ここでα-ケトグルタル酸の2つのカルボキシル基がともに、多分1/2の確率で取り込まれた二酸化炭素に由来するように描いてある。そうすると、このような分布の原因となる段階は2つに絞られる。
1つの可能性はアセチルCoAのオギザロ酢酸のカルボニル基への付加がre面 si面を区別せずに起こる場合であり、いまひとつの可能性はクエン酸からcis-アコニット酸への変換がプロキラリティーを無視して起こる場合である。ところが、最初の付加反応はカルボニル基のsi面側から起こることが知られているし《Biochemistry, 29, 2213-2219(1990)》、クエン酸からcis-アコニット酸への変換は ProR側の側鎖に二重結合が形成されることが分かっている《Proc. Natl. Acad. Sci., 93, 13699-13703 (1996)》。そうなるとα-ケトグルタル酸の2つのカルボキシル基がともに取り込まれた二酸化炭素に由来すると描いてあるこの図はどこかおかしい。さらに、最初に固定された二酸化炭素に由来する、α-ケトグルタル酸のα-ケト酸側のカルボキシル基は脱炭酸で除かれるため、図9のように回路を右回りしてオキサロ酢酸に戻った後アミノ酸への変換が行われるとすれば、そこで生成するアミノ酸には取り込まれないはずである。(図-10参照)
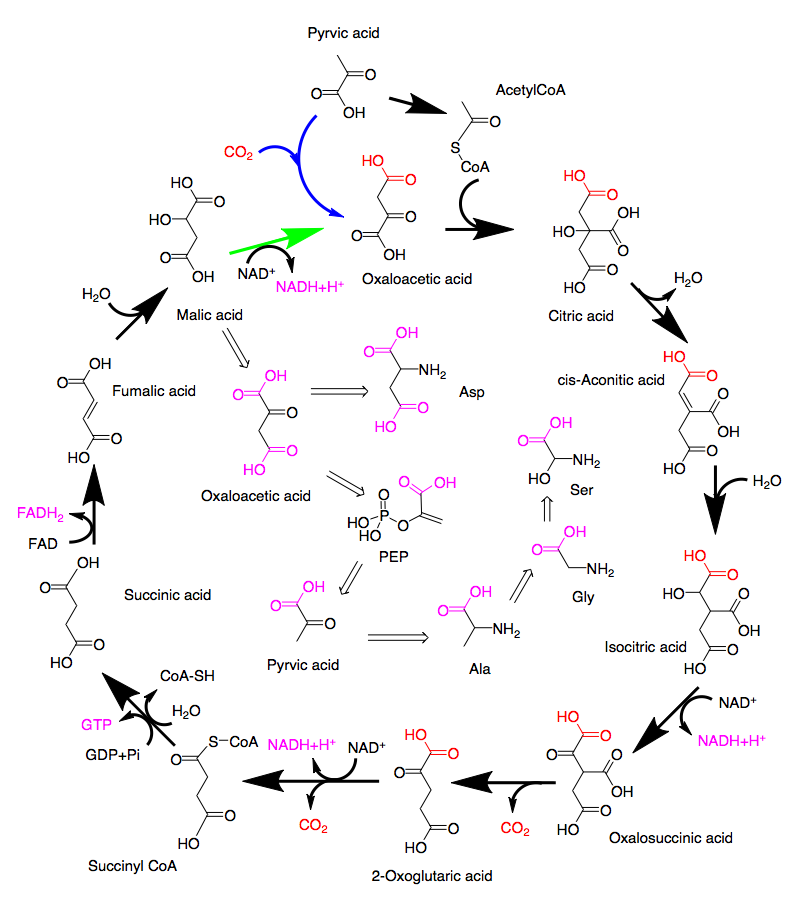
どう考えればよいか?1つの可能性はカイコの持つpyrvate carboxylase [EC: 6.4.1.1]やisocitrate dehydrogenase (EC:1.1.1.41) 、isocitrate dehydrogenase [EC:1.1.1.42]が、プロキラリティを区別する特異性を持っていないと考えることである。そうであれば問題はないが、そんなことはないと思う。この取り込みを説明できる合理的な考え方があり得ると考えている。
公開の予告をしておきながら図-10の描画で戸惑っています。ここをうまく描いておかないと立体化学に精通した人でないと何を言っているのか理解が難しいと思う。プロキラリティの定義を含め、この部分については数日中にアップする予定です。あまり時間が空いても少々寂しいので、取り急ぎ、ここまでをアップしておきます。
歴史生物学 TCA回路への異論 6 に続く