
もうよかろう、言いたい事は言ったか、腹膨るる想いは治まったかと自問すると、まだまだという答えが帰ってくる。確かにフェニルアラニンからリグニンまでの道を辿ることはできた。しかし、植物は何故リグニンを創ったのかという問いには答えていない。世間では、リグニンは植物の体制を維持する役割を持つ、つまりリグニンのおかげで植物は立ち上がることができたというが、些か以上に苦しそうな説明である。何しろ年に200億トンを超える生産量を誇る物質でありながら、植物における存在意義については一行に足りない説明ですむポリマーという点で際立っているわけだ。そうでもないか、デンプンやいくつかの多糖類、あるいはテルペンの一種である天然ゴム(cis−1,4-ポリイソプレン)なども、似たような位置づけにある。リグニンの存在意義について、いくぶん長くなりそうだが、いくつかの事典類がどう説明しているか少しばかり引用する。
日本大百科全書(ニッポニカ)「リグニン」の解説
リグニン lignin
木質素ともいう。維管束植物の道管、仮道管などの木部に多量にみいだされる高分子物質。とくに木材中には乾物量の20~30%に達する量が含まれる。化学構造はベンゼン環に炭素3個がついたフェニルプロパン型の炭素骨格からなり、これが多数互いに側鎖と側鎖、ベンゼン環と側鎖の間で結合した樹枝状構造をもち、分子量5万以上の重合体である。多くの溶媒に不溶のため、パルプ製造の際には亜硫酸処理によってリグニンを可溶化して除去する。
高等植物の分化した組織を薄片にしてフロログルシン(トリオシキベンゼン)塩酸溶液で染色すると、顕微鏡下で細胞壁に沿って赤い呈色が見られる。これはリグニンのコニフェリルアルデヒド基に基づく反応である。細胞分裂を終えたばかりの若い植物細胞は細胞壁もまだ薄く柔らかであるが、しだいにセルロースの一次壁がつくられ固さを増してくる。木化はこのような成長のごく初期からおこり、成長点にある若い細胞でも分裂の数日後にはリグニンの反応が認められるようになる。木化が進むと細胞は分裂の能力を失い、老化して細胞の働きも停止する。しかし、こうして木化が進むことによって細胞どうしが互いに結合し、木部の組織が強固になって、自然の風雨にさらされる植物体をしっかりと保つようになる。細胞壁中ではリグニンはセルロース、ヘミセルロース、ペクチンのような多糖類と強く結合して細胞壁の物理的な強化に役だっている。またセルロースなどの多糖類はリグニンに覆われることで化学的な抵抗力も増す。植物細胞の外側には、初めセルロースを主体とした一次細胞壁が形成されるが、細胞が成熟するにしたがって、セルロースのミセル構造の間にリグニンが沈着し、細胞壁が強固になる。これとともに物質の交換が妨げられ、リグニンが細胞壁全体に広がって一定量に達すると、細胞壁の肥厚が止まる。この過程を木化とよび、あらゆる高等植物細胞でみられる。木化は成長のごく初期からおこるが、木化の進行とともに細胞の老化が進み、最終的には生活力を失う。
植物の進化の程度にしたがい、リグニンの構成成分にいくらかの違いがみられる。裸子植物の針葉樹のリグニンはグワヤシルプロパンがおもな構成単位である。シダ植物のリグニンも針葉樹に似るといわれる。これに対して広葉樹で代表される双子葉植物のリグニンは、グワヤシルプロパンに加えてシリンギルプロパンが主体となり、それゆえメトキシル基CH3O-の割合が針葉樹リグニンよりも高くなる。単子葉植物のリグニンはグワヤシルプロパンとシリンギルプロパンのほかにp-ヒドロキシフェニルプロパンを含んでいる。これらのフェニルプロパン単位が互いに側鎖と側鎖、ベンゼン環と側鎖の間でエーテル結合し、またベンゼン環どうしがジフェニル(ビフェニル)結合して、たいへん入り組んだ重合体となり、リグニンを形成している。
リグニンの生合成は、フェニルアラニンより桂皮(けいひ)酸を経て生成するフェニルプロパノイドが前駆物質となって、これが細胞壁でペルオキシダーゼによって酸化重合して進行する。工業的には、パルプ製造過程における亜硫酸パルプ、ソーダパルプの廃液から大量に回収され、そのアルカリ分解によって得られるバニリンは香料などの工業原料として用いられる。
化学辞典 第2版「リグニン」の解説
リグニン lignin
木材,ワラなど木化した植物の細胞壁の主成分の一つで,木材には20~35%,草本類には15~25% 含まれており,セルロースについで豊富に存在する天然有機物資源である.木材のもつ硬さや,耐朽性,耐候性などの性質は,リグニンの存在と密接に関連している.コニフェリルアルコールのようなフェニルプロパン骨格をもつ構成単位が,ペルオキシダーゼの作用により,無作為に重合したものと考えられる複雑な構造の網目状高分子である.植物により芳香核の置換基が異なり,針葉樹ではグアイアシル核が95% 以上,広葉樹ではグアイアシル核より多くのシリンギル核がほぼ等量からなり,ごく少量のp-ヒドロキシフェニル核も含まれる.禾(か)本科ではp-ヒドロキシフェニル核の存在が知られている.構成単位間の結合様式には,β-O-4型(a),β-5型(b),β-β型(c),β-1型,5-5型などが知られ,(a)がもっとも多い.
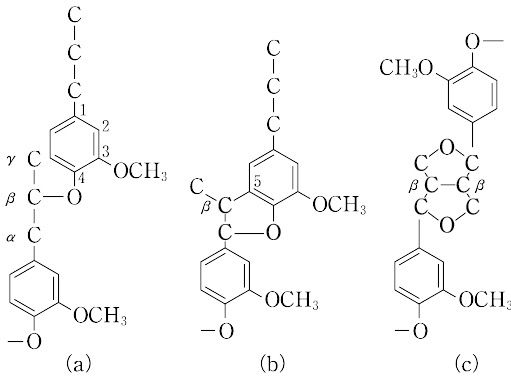
その化学構造の多様性のために単離したリグニンは,きわめて不均質で光学不活性であるが,フロログルシノール-塩酸により特異な赤紫色を呈する。通常,多糖と結合し,リグニン・糖複合体(LCC)として存在する.製紙用パルプの製造では,化学的処理によって植物細胞壁からリグニンを除去することが行われ,これを蒸解という。
Wikipedia Wikipedia からの全文引用では文書量が大きくなり過ぎるので、相当する項目だけを引用する。
存在
リグニンは木材中の20%–30%を占めており、高等植物では生育に伴い、道管・仮道管・繊維などの組織でリグニンが生産される。生産されたリグニンはヘミセルロースと同じくセルロースミクロフィブリルに付着していく。まず細胞間層で堆積が始まり、徐々に一次壁、二次壁へと沈着する。同時にヘミセルロースも堆積し、木化してゆく。構造はランダムでアモルファスである。木部の組織は細胞壁成分だけ残存してほとんどの細胞は死細胞となり、通導・植物体支持を担う。腐朽・食害への抵抗性を有する。
食品では、亜麻仁(フラックスシード)、根菜類(ニンジン、パセリ、ダイコン)、小麦ふすま(wheat bran)などに比較的多く含まれるとされる。
ボールドで表示した部分に取って付けたかの如くリグニンの機能が書いてあるが、植物は何故リグニンを創ったのかと言う問いかけはない。創り出されたリグニンに、書いてあるような機能はあるだろう。しかしそれは創り出された後の話である。植物がリグニンに書いてあるような機能があることを予知して、あの複雑な生合成系を植物が作り上げたはずはない。歴史生物学的視座に立てば、植物は何らかの必然性を持ってリグニンを創った。その結果、創り出されたリグニンが述べられている機能を持っていたという時系列の物語になるはずである。ではその「何らかの必然性」とは何かというのが次のテーマになる。
偉そうに書いているけれども、実はこの程度の事は今までの研究者も知っていたことである。彼らは賢かったため、答えにくいことに蓋をしていただけだとと思う。時々というか、まあ世間の常識を知りたいと思ったら、Wikipedia を参照することが多いのだが(信じることとは違う、でも有り難いと思う場面は多い)、適応という項目の中に蓋をしていることを示唆する文章がある。
「適応と目的論
通常、生物学者は適応的な形質を「○○のための形質」と呼ぶ。このために、しばしば進化には意図や方向性がある、または目的論を含意していると誤解される。このような表現は「自然選択によってその形質に影響を与える一連の遺伝的変異が蓄積され、その形質が形成された」と言う表現の短縮形である。」
そうなんだ。目的論は含んでいないんだ。無知無学で世間知をに欠けた私が、皆様の暗黙の了解を阿呆臭く突いていただけの事ということらしい。でもね〜、例えば、イモガイの持つコノトキシン、説明は「この貝は毒の入った銛のような歯舌で小さな魚を刺し、麻痺させて捕らえる。」(http://www.sc.fukuoka-u.ac.jp/~bc1/Biochem/venoms.htmより) と書いてある。この文章を、「イモガイにおいては自然選択によってタンパク質毒素コノトキシンを作らせる一連の遺伝的変異が蓄積され、そのタンパク質毒素コノトキシン生合成能が形成された。」という表現の短縮形として読めというわけだ。上記のサイト、こんな形で引用するのは気が咎める。ごめんなさい。
上記のhttp://www.sc.fukuoka-u.ac.jp/~bc1/Biochem/index1.htm の各ページにおいては、丁寧な図と説明がなされており、初学者には有益なサイトである。私も時々訪れて、知識の確認だけでなく、知らなかったことを学んでいる。ちょっとばかり恩を仇で返したような気分である。もちろん、時にはこのサイトの作成者と私の間で視座の違いによる意見の違いが発生するが、それは仕方がない。それにしても上記の短縮形という意見を認めるとすれば、NHKラジオの「夏休み子供相談」なんて、聞いてられないな。
「適応主義
生物の形質や行動はほぼすべて適応的であると仮定して理論を構築する立場を適応主義という。適応主義は現在、批判的な意味と肯定的な意味で異なる意味で用いられる。肯定的な意味では「生物はなぜそのような機能的な特徴を持っているのか?」という疑問のことである。その答えが「たまたま偶然」だったとしても、なぜ?と言う最初の疑問は正当である。批判的な意味では、十分な証拠無しに生物の特徴を適応であると結論する研究や研究者のことを指すことが多い。」
次の項目は適応主義についての説明だが、ここには私の持つ疑問を肯定するフレーズがある。「生物はなぜそのような機能的な特徴を持っているのか?」という疑問のことである。その答えが「たまたま偶然」だったとしても、なぜ?と言う最初の疑問は正当である。そう、このブログでは、その正当な疑問に正面から答えようとする意図を持った説明が、アブシジン酸や、花の色素や、アルカロイドなど殆どの植物成分に対して全く行われていなかったことに対する細やかな抵抗である。
とすれば、リグニンの raison d’être 1 は、リグニン生合成研究の歴史をなぞったにすぎない。いままで述べたことは、単に現生植物の生合成経路解明の歴史であり、このブログの目的からすれば前書きに対応する。それにしては長かったなという思いは在るが、ここから、どのような本論がが可能になるのだろう。
歴史を重視する歴史生物学を標榜する人間として、まずリグニンがいつ頃から作られ始めたかという疑問は避けて通れない。リグニンはいつ頃から地球上に存在するか?大まかな答えは得られている。植物化石に存在する細胞壁の肥厚組織をリグニンの存在と結びつけて、シルル紀からデボン紀頃のリニア類植物にはすでに存在したとする判断である。デボン紀は4.16億年前から3.59億年前の時代を意味するが、この時代の気候は温暖で、最古の木であるアーケオプテルス(Archaeopteris)が繁茂する最古の森林が存在していたことが分かっている。この木は地球上に初めて現れたシダ植物の仲間の木本植物で樹高30 mにも達したらしい。

樹高が 30 mもあったとすれば、強度の観点から見て当然リグニンを含んでいたに違いないと推測される。従ってリグニンの起源はそれより前の時代に求めなければならない。シルル紀はおよそ4.37億年前に始まり4.16億年前までの期間を云うが、陸上植物としてすでに立ち上がったクックソニアの化石が4.1億年前の地層から得られている。これを考慮すれば、植物の上陸はもう少し前のオルドビス紀まで戻る可能性が高い。コケ類の胞子と推定されている微化石が、4.7億年前の地層から発見されており、植物の上陸はこれに先行すると考えられるからである。まあ具体的な証拠はないのだが、植物が陸上へと進出するに際して、生合成が始まったと書いてある場合も多い。こういう事象群を総合的に判断すると、リグニンの生合成はどうやらオルドビス紀(約4億8830万年前から約4億4370万年前まで)において開始された推測して良いだろう。
さて、だらだらと記述を続けてきながら、上記の部分がリグニンの歴史について述べた本論です、はいおしまいではバランスが悪すぎる。ここで終わるのでは余りにも芸がないと思う。とはいえ、タイムマシンはないのだから、現場を見ることはできない。これから議論を発展させるとすれば、空想と妄想の織りなす世界を、可能な限り理性的に切り分けていくしかない。議論が合理的であるかどうかの判断は、古生物学的知見と生化学的知見、そして地球化学的な知見をアンカーして使うことにする。ではここからいかなる議論が可能になるのか。
原点に戻ろう。私の持った根源的疑問は、「なぜ植物はリグニンを持つか、あるいは作るか?」という点にある。科学に持ち込んではならないと教えられた「何故・Why」と言う問いである。長々と述べてきたことは、どのように作られるかという「How」に対する答えであった。[How]に対する答えは科学という土俵に乗るが、「Why」に対する答えは物語になる。物語は科学ではない。現役時代の私が一番悩んだ点である。いまは無役、言い換えれば自由である。自由の代償として、間違いなく貧乏ではあるが、この自由さを利用して想像の翼を目一杯に広げ、大きなそして少しだけ裏付けのある法螺を吹ふいてみることにしよう。
何度も云うようだが、リグニンは木材の20~30%を占める極めて安定な高分子である。いままでは、単にリグニンと呼んできたが、リグニンにも種類がある。リグニンは、主としてイネ科植物に存在する草本リグニン、針葉樹に分布するグアイアシルリグニン そして広葉樹に分布するシリンギルリグニンに分類されている。各リグニンに対しては膨大な研究が行われており、かなりなことが分かってきたと考えて良い。
さて、植物においてリグニンはセルロースと一緒に存在する。リグニンの生合成は5億年より少し後に始まっただろうと推測したが、セルロースはいつ頃から作られたのだろう。そういう立場から眺めた人はあまりいないようだが、石炭は植物のリグニンやセルロースに由来するという記述がある。さらに、バクテリアは bacterial cellulose をつくるとともに葉緑体の祖先であるシアノバクテリアの仲間もセルロース生合成系を持つことを考えれば、セルロースの方がリグニンより歴史は古そうである。ただ、石炭の化学構造の例を見る限りにおいては、石炭はリグニンが主原料であると思う。

さて、セルロースの分解酵素はかなり広い生物中に存在するが、リグニンの分解酵素は生物は白色腐朽菌であるシイタケ、ヒラタケ、マイタケの仲間しか持っていない。これは歴史の長さを反映した結果なのだろうか。
2012年にリグニンを分解するペルオキシダーゼのアミノ酸配列の解析から、白色腐朽菌がリグニン分解能を獲得したのは石炭紀の終わり頃であるとする報告が出された。The Paleozoic origin of enzymatic mechanisms for decay of lignin reconstructed using 31 fungal genomes; Science 336, 1715 (2012) 著者は70人の連名であるため省くことにするが、彼らは31種類の真菌類ゲノム配列に対して分子時計解析を行い、担子菌門からハラタケ亜門が分かれたのは約4億7千万年~4億3千万年前で、ハラタケ亜門からリグニン分解能力を初めて獲得したハラタケ綱が分かれたのは約2億9千万年前と推定した。また同様の解析をペルオキシダーゼのアミノ酸配列に対して行い、リグニン分解能を持つマンガンペルオキシダーゼ遺伝子は2億9千5百万年前に出現したと推定している。これらの結果は、最も古い白色腐朽菌の化石がこれまでペルム紀(約2億6千万年前)から見つかっていること、さらに石炭紀からペルム紀への移行期に急速な有機炭素貯蔵量の減少が起こっていることの説明になると結論している。納得できる結論である。
そうか、古生代のデボン紀から石炭紀にかけてのシダ植物を主要メンバーとする大森林時代においては、倒れた木のセルロースは分解されるにしてもリグニンはひたすら貯まっていくだけだったのである。少し考えれば分かることだが、デボン紀は4.16億年前に始まる。先に述べたアーケオプテリスはデボン紀の植物である。これに続く石炭紀は3.59億年ほど前に始まり2.99億年前まで続くわけである。この間、約1億年、この1億年の間にひたすら貯まり続けた木材が地殻変動により地中へと引き込まれて石炭化する。そうであれば、この時代に貯まりに貯まったリグニンが炭化した結果としての石炭だとすれば、その膨大な存在量が納得できる。
いままでの説明を総括すると、水中にいた植物が陸上への進出に当たって、体を支えるための支持機構を必要とした。この目的にかなう植物成分がリグニンである。植物はセルロースを鉄筋、リグニンをコンクリートとして陸上環境に適応し、デボン紀から石炭紀にかけて、陸上に大森林を形成した。セルロースに関しては、セルラーゼを持った生物がすでにいたようで、どの程度の量が残ったか分からないが、リグニン分解能を持つ生物は2.95億年に出現する白色腐朽菌まで存在しなかった。従って、大森林時代の植物の遺骸に含まれるリグニンはほとんど分解されることなく堆積し、これが我々がいま利用している石炭(無煙炭)の原料となった。ということになるのだろう。
多くの読者は上記の総括で納得してくれるだろう。だが、上の文章は、私の意見を押し殺して、一般的知識人の発言するとすればこうなるだろうと予想して書いたものである。自ら書いた文章にケチをつけるのは矛盾しているが、上の文章を批判することにする。
いつものことだが、この手の書き方には納得できない。なぜリグニンが生合成されたかという問題を実に巧妙に回避しているからだ。この何故は、一つ間違えると科学の領域を超えた議論に導かれる危険なモノであることは承知している。しかし、余りにも無造作にこの問いから逃避することは、科学の深化にとって有害であると考える。いくつかの知見を組み合わせて、熟考してみたい。
まず、酸素は毒物である。種々の要因の存在下に酸素は活性化され、反応性の高い活性酸素と呼ばれる分子種になる。この分子種には一重項酸素、ヒドロキシラジカル、過酸化水素、スーパーオキシドアニオンラジカルが含まれる。光合成を行う植物細胞中には、光で励起されたクロロフィルがいわゆる炭酸同化で使い切れなかったエネルギーを、近くにある酸素分子に供与することで発生する。活性酸素は毒物であることから、植物細胞にはSOD(スーパーオキシドディスムターゼ)、ペルオキシダーゼ、カタラーゼなどからなる酵素的消去機構が存在するだけでなく、β-カロテンを初めとするカロテノイド、トコフェロール(ビタミンE類)、アスコルビン酸(ビタミンC)、グルタチオン、フラボノイドなどの低分子物質が活性酸素クエンチャーとして存在し、その毒性から回避している。だから、植物成分に含まれる抗酸化物質を食べて老化を防止するなどという言説が成立しているのだが、健康のためにはその辺にある旬の野菜を食べるだけで十分であり、無理に偏食ともいえるサプリメントを求める必要はない。
さて、植物が上陸した5 億年ほど前の地球を見てみると、23 億年頃に 3%程度まで急上昇した酸素濃度は、6.4 億年頃には再度上昇し5 億年前頃は 12~13%まで上昇していたと考えられている。その結果何が起こっていたかと云えば、オゾン層の形成が起こっていたのである。酸素分子は242 nm以下の短波長紫外線を吸収して解離し、生成した酸素原子は近辺の酸素分子と反応してオゾンを形成する。オゾン分子は320 nmよりも波長の短い紫外線を吸収して酸素分子と酸素原子に分解する。何だ、何も変わらないではないかと思われるかもしれないが、紫外線の持つエネルギーはこの 2 つの反応を通して熱エネルギーに変換されている。その結果、オゾン層あたりの温度はかなり高い。とにかくオゾン層の成立に伴い、太陽光に含まれるUV-C (200~280 nm)は完全に吸収され、UV-B(315~280 nm)の 99.5%とUV-A (400–315 nm)の94.5%はオゾン層で遮られる。オゾン層の成立という条件があったが故に、植物の上陸は可能になったといえる。
植物の立場から上陸を眺めると、もう一つ重要な壁があったと思われる。当時の大気中における酸素濃度が 13%であったならば、この大気と接する水系の酸素濃度はどれくらいだっただろう。現在の大気組成で 25℃における酸素の飽和溶存量はおよそ8 ppmである。現在の酸素濃度を21%、当時の酸素濃度を13%とすれば、当時の25℃における水系の飽和溶存酸素量は 5 ppm 程度だろう。もちろん、溶け込んでいる塩類の量と組成でこの量は変動する。光合成をしている植物が繁茂しているところでは過飽和になる場所があったと思うし、藻類の細胞内酸素濃度はもっと高かった可能性は十分にあるわけだ。そうした酸素濃度の高いところで、酸素毒性の消去能を強化したグループが上陸を始めたに違いない。とは思うが、空気中の酸素濃度は水中の酸素濃度の 30 倍を軽く超える。植物のパイオニア達は、酸素発生型光合成を持って 30 倍以上の酸素濃度を持つ大気中に進出を試み、これに成功したわけである。
「多糖類の存在意義 」 の中で述べる予定だが、植物の上陸は海ではなく、陸封化された巨大の塩水湖が次第に淡水化した岸辺から起こったのではないかと考えている。上陸に際して、浸透圧、酸素濃度、乾燥、温度変化、重力、光強度などの多くの否定的条件を一気に克服するのは無理だろうと考えたわけだ。温暖な淡水湖から湿地帯を経由して上がれば、浸透圧、乾燥、温度変化の条件は除外できることになる。残った、酸素濃度、重力、光強度には通底する問題が含まれているようだ。考えて欲しい。海岸には植物は殆ど生えていない。海から上陸したのであれば、海岸は母なる土地であり、植物は繁茂しているはずではないか。現実の海岸は、世界中どこでも植物のない砂浜や岸壁が続いている。
こんな文章を書いていて困ってしまうのは、全体のデッサンなしに書いていることに起因する。大きな枠組みについての朧気なイメージはあるのだが、個々の事項をどこで扱うかについては、何のイメージもない。しまった、あそこで書きすぎたとか、これからどう続ければよいかなどと悩むことばかりである。もちろん書いたことはないが、なんだか新聞小説を書いているような気分である。作家の人たちは、どのように全体の構成を考えているのだろう。
現代という時代の底流には、全てのものに対してそれはお金すなわち利益を生むのかという判断基準があるようだ。好きではないが、その価値観が日々強くなっているのは間違いないだろう。お金に縛られないとされていた学者(研究者)と呼ばれる人たちでも、この研究が3年あるいは5年後に終わると、社会に対してこのようなインパクトがありますと申請書に書かねばならないご時世である。インパクトとは商売のチャンス、あるいは萌芽と言い換えてもいい。あらゆることにコストパフォーマンスを求めるのが現代の社会らしい。
ところがである、植物がある化合物を作る、例えばリグニンを作るという場合、その生合成にどれほどのエネルギーが必要であるのか、そのエネルギーの消費に対して、作られる過程、あるいは作られた化合物はそれに見合う効果を示すのかという視座からの研究は極めて少ないようだ。私は寡聞にして一つの報告しか見たことがない。コストパフォーマンスで縛られている研究者が増えている現在、もしカール・マンハイムの言説が正しければそのような研究が行われても良いような気がするのだが、若い研究者は初めからそのような環境で育ったために、コストパフォーマンスに縛られているという感覚そのものがないのかもしれない。やりたいことだけをやってきた私達の世代の方が、そうした強く束縛を感じるのであろう。
そういえば、学園紛争華やかりし頃に、産学協同は唾棄すべき悪であると叫んでいた人達はどうしているのだろう。皆大人に、従順な社会人になったのだろう。当時私は、自らの主体性を見失うことがない程度の産学共同はあり得ると考えていた。それ故に一部の人達からはお前は右翼だとの批判を浴びた。だがその気持ちはいまも全く変わっていない。いまは時々左翼だと位置づけられる。周りの人たちが、私の右に行ったり左に行ったりしているだけのような気がしている。それ以上に、こうした問題自体が、右・左で捉えるものではないと思う。それはそうとして、あの時代を生きた旧世代の人のほうが、束縛の中にいるという視座を持ちうる蓋然性が高いのではないか。実はいまから、そういう視座からの話をしたいと思っている。
リグニンの生合成系、図 29を思い出して欲しい。いやここに再度示す事にしよう。
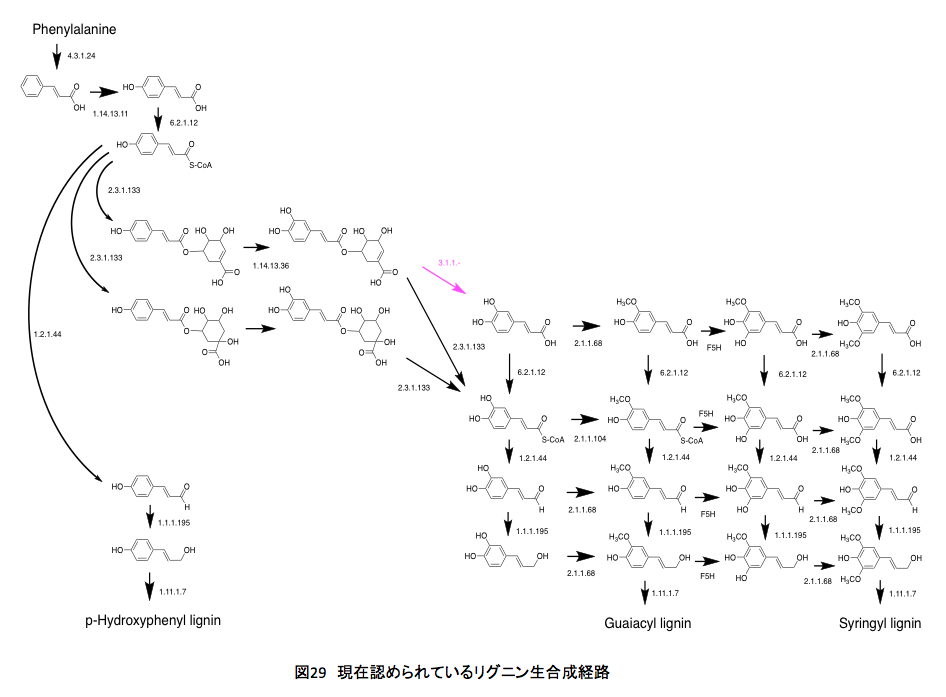
この図には、リグニンの構成成分となるフェニルプロパンユニットがどのように作られてくるかを、外部から関与する物質抜きで描かれている。さてこの図に描いてある各段階の反応に、それぞれの段階において消費される高エネルギー物質を付け加えると図 32になる。いやはや、結構面倒な作業であった。
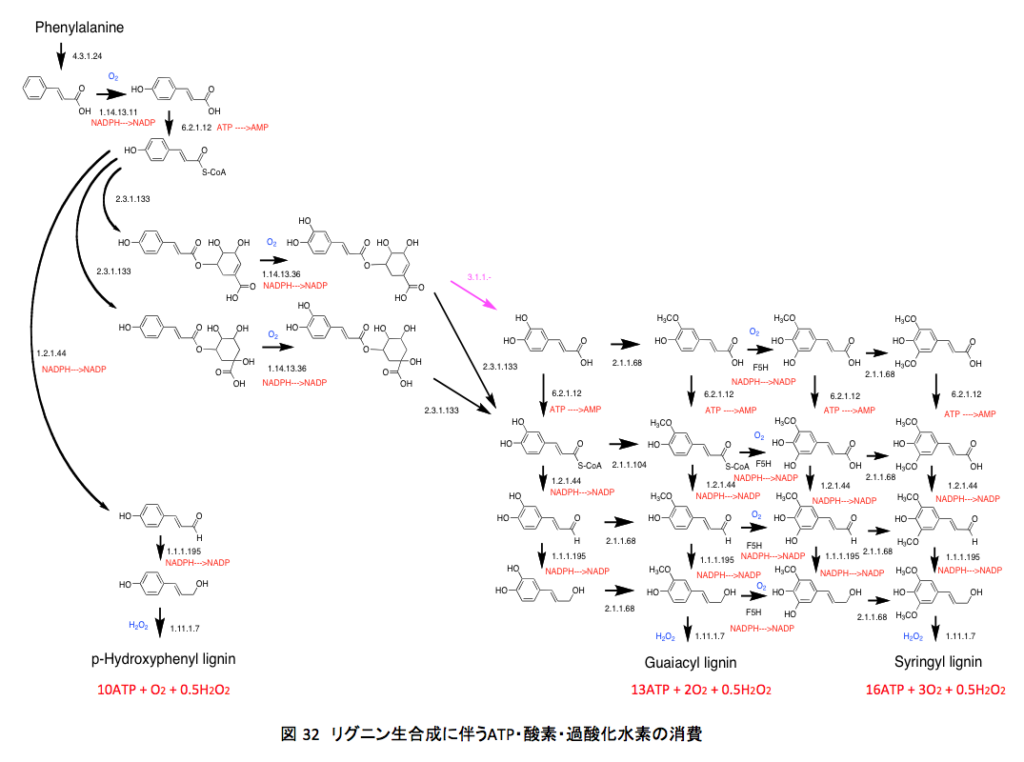
最下段に、フェニルアラニンからリグニンまで誘導されたとき、フェニルプロパン1ユニット当たりで消費されるATPと酸素分子と過酸化水素の分子数を示している。本来ならアセチルCoAからフェニルアラニンまでのATP、NAD(P)Hの消費や、S-アデノシルメチオニンの代謝で使われるATP、NAD(P)Hの消費を精査すべきであろう。ただ、そこまでやろうとするとかなり大変な作業となってしまい、収拾がつかなくなる可能性と同時に、途中で間違ってしまう可能性が極めて高い。従って、ここでは論旨を説明するのに必要且つ十分な範囲内での計算としておく。上記の範囲であってもどこかにミスがあるかもしれない。消費するATPの計算において、NADPHのNADPへの変換は、電子伝達系を通った場合3ATPが生産されることから3ATPと、ATP—>AMP+PPへの変換は2ATPの消費としてみてもいいのだが生産物であるピロリン酸にまだエネルギーが残っていると考え1ATPの消費であるとして計算した。
そうするとp-ヒドロキシリグニンまでの1ユニット当たりのATPと酸素、そして過酸化水素の消費量は10 ATP、1 O2、そして0.5 H2O2となる。同様にグアイアシルリグニン、シリンギルリグニンに対しては、それぞれ13 ATP、2 O2、0.5 H2O2、そして16 ATP、3 O2、0.5 H2O2という値が得られる。少しばかり身近な量に換算するが、1トンのp-ヒドロキシリグニンが作られるとき、消費されるATPは 33.8トン、酸素は 213 Kg、過酸化水素は 119 Kg、1トンのグアイアシルリグニンが作られるとき、消費されるATPは 33.8トン、酸素は 213 Kg、過酸化水素は 119 Kg、シリンギルリグニンであれば消費されるATPは 47.3トン、酸素は 639 Kg、過酸化水素は 119 Kgとなる。1トン当たりで計算したが、リグニンの年間生産量は200億トンを超えると推定されている事を考慮すれば、その量の多さは絶句するほどだ。
これらの系におけるATPを消費量には驚かされるが、これはあまり気にしなくて良さそうだ。消費されたからといって消えてなくなるわけではない。使用済みのADPやAMPは光合成系やグリコリシスにおいてすぐにATPへと再変換される。従って、実際に動いている量は遥かに少量であることが予想される。重要なのは思いもかけないほど多量に消費される酸素と、過酸化水素の方だろう。過酸化水素に関しては、この物質が活性酸素であることから、いわゆる活性酸素消去システムによって消去されたと考えればよい。では酸素をどう考えればよいか。筆者はこれは最も重要なことだと考えている。何故か? 改めて述べるのは幾分億劫であるし、記憶力のいい人にとっては鬱陶しいだけだろう。CYPの役割についての仮説を https://bamboolab.yamasatoagr.com/?p=6420 と https://bamboolab.yamasatoagr.com/?p=6422 に書いている。
植物は上陸に際して、強い光と水中に較べて30倍以上の酸素濃度を持つ大気中に進出を試みたわけである。光合成色素を持つ(これは光増感剤しても働きうる)植物が上陸したときに、まず遭遇する厳しい障壁がこの二つであったと考えて良い。生成する活性酸素の毒性を消去するためには、スーパーオキシドアニオンラジカルの処理が必要なのだが、困ったことにスーパーオキシドディスムターゼによる解毒処理に伴って、酸素分子が再生するだけでなく別の活性酸素分子種である過酸化水素が生成する。過酸化水素にはカタラーゼが働くのだが、困ったことに酸素分子が再生してしまう。
スーパーオキシドアニオンラジカルは消去系と名付けられてる上記の反応系では、スーパーオキシドアニオンラジカルと途中に生成する過酸化水素は消去するが、スーパーオキシドアニオンラジカルの原料である酸素分子を再生してしまう。植物としては細胞中の酸素濃度を消去することができればより好ましいのではないか。強い光が原因で生合成しすぎた糖については、多糖として系外に追い出すか光呼吸系を通して、もう一度二酸化炭素に戻せばよい。などという種々の反応などを総合的にあれこれ考えていると、リグニン生合成系は活性酸素の原料である酸素分子その物を消去することで、酸素毒性回避を行う系であると考えることができると思う。
植物は光合成で作りすぎたATPを消費し、酸素分子をクエンチし、過酸化水素を消去するリグニン生合成系を獲得した結果、重力へ対応するのに極めて有益なリグニンという高分子物質をも獲得したのである。一旦、獲得したリグニン生合成系を、陸上植物が決して失わなかったのは上記の理由からではなかったか。リグニンは植物の体制を維持するために作られたのではなかった。細胞内に酸素発生装置である葉緑体を組み込んだ植物は、基本的に酸素過剰という矛盾を内包した生き物である。リグニン生合成系はこの過剰な酸素を消費する系としてこれを獲得した。その結果多量に生産されてくるリグニンをセルロースやヘミセルロースミクロフィブリルに付着させ、細胞質の系外に追い出したのである。その結果として細胞璧は強固になり、植物の立ち上がりを可能にしたのみならず、1000年を超える風雪に耐える強固な体制の構築を可能にしたのである。
最後に一言、植物と動物は違う生き物である。動物は車に似ている。燃料(ATP)が満タンであるときが最も元気なときであると考えて良い。しかし、この考えは植物に対しては通用しない。光が当たって光合成が動き始めたときにATPしかなかったら、光リン酸化がうまくゆかず致死的な傷害がおこる。植物の生存にとっては適切なATP/ADP 比の維持が不可欠であり、ATP/ADP比が一定以上大きくなることは命にかかわる異常事態なのである。
終章の部分には今後書くべき多くの課題が存在する。一つ一つに拘泥すると、内容は凄まじい量になってしまう。従って、リグニンのraison d’être (存在意義)については、ここで一旦筆を置くことにする。リグニンのraison d’être1 よりもリグニンのraison d’être 2 の方が面白いと感じた方のほうが多いのではないだろうか。
もう午前4時、今日か明日かわからないが朝刊がきた。昼間はなかなかお金にならない農作業で疲れているのだが、毎日、朝刊を運んでくる50 cc のバイクの音を聞いてから寝るのが日課になった。少しだけ反省しよう。