
1945年にホロウィッツは前生物的に生成していたアミノ酸、糖、核酸塩基、脂質などが蓄積していたことを前提に、発生した原初生物がこれらを基質として利用していたとする条件下に、代謝経路は最終産物から中心代謝へ向かって逆向きに伸長したとする説を提唱している。つまり生存に必要な基質の不足を、その基質の前駆体に求めることで代謝系が創生されていったとしたわけだ。この説に従えば、プレビオティックに作られていたグルコースが、その利用系が創成されるまで大量に蓄積していたことを意味する。しかし、そんな予定調和みたいな物語で説明の付くことではないだろう。
さて、宇宙空間には星間分子と呼ばれる一群の化合物群が分布している。詳しくは専門書を参照して欲しいが(http://www.astrochemistry.net/ )にもある程度詳細なデータが記載されている)、700種を越える発見された星間物質の中には、ホルムアルデヒド、アンモニア、シアン化水素、ギ酸、メタノール、ビニルアルコール、アセトアルデヒド、グリコールアルデヒド、酢酸、エタノール、アセトン、ベンゼン、ナフタレンなどとともに、グリシンやジヒドロキシアセトンなどが見つかっている。William M. Irvineによれば、銀河系に存在するエタノール量は8x1024トンに上るという。とても飲みきれない量である。確かに広大な宇宙空間には、こうした化合物群が生物の関与なしに存在しているとはいえ、それらの存在密度は極めて低い。
さらに、原始地球大気の組成の問題ゆえに、いまとなっては否定的に見られているとはいえ、ユーレイ-ミラーの実験に端を発する非生物的な有機化合物の合成実験で、核酸塩基、アミノ酸をはじめとして、多種の糖(グリコールアルデヒド、グリセルアルデヒド、ジヒドロキシアセトンなど)や低級脂肪酸類(ギ酸、酢酸、プロパン酸、乳酸、ピルビン酸など)が比較的容易に生成することが明らかになっている。そして、星間物質としても知られるホルムアルデヒドは、粘土鉱物上で連続的に縮合してホルモースと呼ばれる多種の糖の混合物を与えるのだが、この混合物の中に5単糖、6単糖が含まれる場合があると報告されてはいる。しかしながら、α-D-グルコースという6単糖の1光学異性体だけが、特異的に大量に蓄積する可能性は極めて低いだろう。初めて出現した生物が従属栄養生物であったとして、彼らの出現時に生存と増殖を担保するような量のグルコースが、前生物的に蓄積していたとする物語は成立しないと考える。
一方、もし原初の生物が独立栄養生物であったならば、原初の解糖系の流れる方向は3単糖から6単糖に向かうベクトルを持ち、グルコ−スは原初生物自らが創生したものとするのが理性的判断ではあるまいか。こう書くと、原初の生物が独立栄養生物であるはずはないとする批判を浴びることがあるのだが、いわゆる独立栄養生物、従属栄養生物という分類は、環境から取り入れる物質の複雑さの程度に差があるだけで、本質的な差はない。いずれに分類される生物であっても、生存に必要な物質を環境に依存していることに変わりはない。さらに原初の生物は解糖系で得られるATPに依存していたのではなく、いわゆる酢酸などのC2化合物や地球化学的に存在するポリリン酸の代謝を含む原始発酵系で得られるATPを使っていたと思われる。
時として、お前は変わっていると評価される。自覚はない。よくよく話を聞いてみると、考える際の空間的・時間的スパンが相互に大きく異なっている場合が多い。私のスパンの上で、私はすこぶる常識的な人間である。その極めて常識的な私が素直に考えるとすれば、原初生物におけるグルコースの生合成は、グルコースの分解系あるいはグルコースの利用系に先立って起こったと思惟する。グルコースの生合成が行われた後に、グルコースの利用系としての解糖系が成立したと考えるべきであろうし、グリコーゲンやデンプンなどを含む多糖類の生合成系が成立した後でそれらの利用系が成立したと考えるべきではないだろうか。尤も、こんな複雑な系が相互に独立して創生されることはなかなか考えにくい。二つの系で多くの段階が共通であり、かつ酵素反応の可逆性を考えれば、生成系と利用系が同時に成立していったとする考え方が極めて理性的であろう。
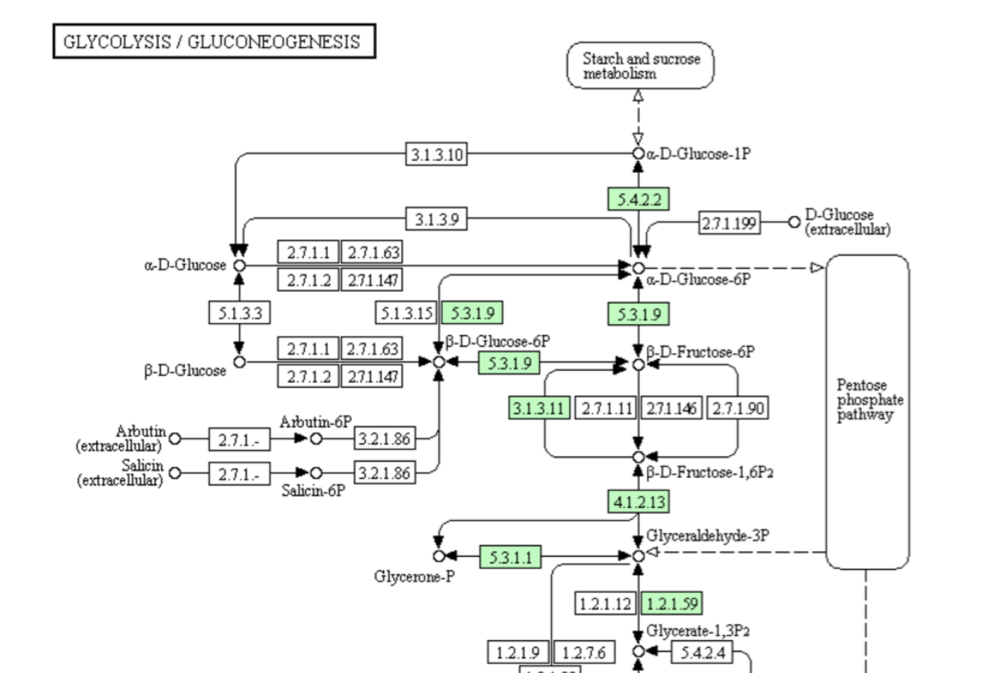
さて、高校および大学初年度までの段階ではほとんど触れられることはないが、解糖系に寄り添う一つの代謝系がある。図9に示す糖新生系路と呼ばれているこの系は、人に植物に大腸菌に、そしてアカパンカビにも古細菌にも分布する。そしてその分布は解糖系以上に広い。

さて、解糖・糖新生系に限らず殆ど全ての代謝系で系の作動を担保しているのは、酵素という触媒機能を持つタンパク質である。この酵素は、触媒としての機能を持ち、反応速度を大きく時には劇的に変化させる。しかし、反応の平衡点を移動させることはない。平衡点は熱力学的に決まるものである。つまり、AからBの化学変化を触媒する酵素は、AとBの濃度比によってはBからAへの変化をも触媒するのである。理論的には、グルコースからピルビン酸への代謝系はピルビン酸からグルコースへの変換も可能であることを意味する。実際の解糖系は、逆反応が起こらない不可逆なプロセスを含むため、そのまま逆行することはできない。しかし、そこには逆行を可能とするバイパスが存在するため、系全体としての逆反応が可能になっている。図9においては、解糖系と異なる段階を点線の矢印で示している。
さて解糖系は、ブドウ糖やブドウ糖ユニットで構成される糖質を栄養源として取り込む従属栄養生物においては解糖を進める方向で機能するが、これに寄り添う糖新生系は、同時に糖を新生する方向で動いているのである。ましてや、そうした栄養源としての糖質に依存しない独立栄養生物においては、糖新生系が先に稼働するのは自明ではないか。
実は幾分以上に、自ら展開している議論に不満である。糖新生系の定義を明確にすることなく議論に入っているからである。通常、糖新生系は解糖系(EM系路)の逆反応とされている。では糖新生系は解糖系の最終物質であるピルビン酸から始まるかと云えばそうではない。ピルビン酸も出発物質の一つとして含まれはするものの、乳酸、プロピオン酸、糖原性アミノ酸などから、おおむね解糖を逆行してα-D-グルコースをつくる経路を糖新生系(Gluconeogenesis)と記述している場合が多い。解糖系においては、乳酸まで含めれば酸化還元のバランスがとれると説明できるのにこれを外し、糖新生においてはピルビン酸から始めればEM系路の逆向きの反応であると云えるのに、ピルビン酸だけでなく乳酸、プロピオン酸、糖原性アミノ酸などを加える。推測だが、筋肉生理をやっていた人たちの考えが、色濃く反映しているように感じられる。
私見だが、この糖新生系は解糖系に比べより根源的で重要な系路であるように思える。にもかかわらず、言及される頻度はあまり高くない。何故か?解糖系が1分子のグルコースから2分子のピルビン酸に至る過程で2分子のATPを生産するのに対し、2分子の乳酸から1分子のグルコースを生産する糖新生系においては6分子のATPが消費されるからであろう。どうやらエネルギーを生産する系の方が、世の中では重要視されているようだ。20世紀から、エネルギー源である石油利権を巡って争い続け、さらには将来のエネルギー源を何に求めるかで混迷を深めている現在の世界状況を考えると、こうしたエネルギー源を重視する社会的な思考の枠組みが、代謝系の意義づけに影を落としてはいるのではないだろうか。
さて、この世には種々多様な生物が棲息している。そのなかに温泉や熱水噴出口、あるいは塩濃度の高い湖に分布する変わった細菌がいる。いまでは古細菌と呼ばれるこの生物は、古くから何かしら変な細菌として一部の研究者には知られていた。16S rRNA配列から全生物の分類を試みたカール・ウーズの野心的な研究が進展した1970代になって、彼らは分類学上で古細菌という新たな分類学的位置を与えられ、広く認知されるようになった。この生物は16S rRNAの塩基配列において真核生物ともバクテリアとも大きく異なるとはいえ、バクテリアより真核生物に近い生物である。
我々の周りにも色々な古細菌が棲息するが、その中で比較的知名度の高い古細菌はメタン細菌であろう。1988年以降、世界を振り回してきた地球温暖化、その一つの原因としてのメタンを発生させる細菌である。地球温暖化仮説が科学理論として成立するかどうかは少々以上に疑わしい所があるとはいえ、メタン菌がメタンを作るのは間違いない。一時は、メタンを含むウシのゲップを止めろとか、メタン発生源である東アジアの稲作を制限しろとか、気違いじみた議論がなされていたのが記憶に新しい。2018年時点で、500種を越える古細菌が知られているが、KEGGには以下に示す281種のデータが記載されている。括弧内は知られている種数に対しブドウ糖合成酵素もブドウ糖リン酸化酵素も持たない種の数を示している。Euryarchaeota (45/191) Nanohaloarchaeota (0/1) Crenarchaeota (34/64) Thaumarchaeota (15/16) Nanoarchaeota (1/1) Micrarchaeota (?/1) Korarchaeota (1/1) Bathyarchaeota (2/2) Lokiarchaeota (0/1) Unclassified Archaea (2/2)
余り世の中をかき回したくはないが、この中にG-6-PやG-1-Pまでの代謝系は持ちながらも、ブドウ糖に変換する酵素とブドウ糖をリン酸化する酵素を持たない種が、ユーリアーキオータ45種(191種)、クレンアーキオータで34種(64種)、タウムアーキオータで15種(16種)、などが存在する。ブドウ糖合成酵素を持たないだけの条件にすれば、ほとんど全ての種が当てはまる。もちろん、Sulfolobus tokodaiiにおいて基質特異性が低くADPをリン酸供与体とするヘキソキナーゼ(6単糖リン酸化酵素)が見つかったように、いくつかの種では起源の違う加水分解酵素やリン酸化酵素を持つ可能性が残るとは云え、全部の種が消えることはないだろう。
こうしたブドウ糖まで延伸していない解糖系は、古細菌にのみ分布しているわけではない。真正細菌《バクテリア》の中にも、超好熱細菌であるAquifex aeolicusだけでなく、Bacteroidetes門に属するかなりな数の微生物に分布している。更に、純粋培養ができていない微生物―分類名の前にCandidatusが付いている−においては、その分布割合はかなり高い。(ただし、これらの微生物の中には他の生物に寄生することで生活している種がかなり含まれており、共生進化の過程で解糖系を失った可能性については、注意を払う必要がありそうだ。)
解糖系と書くと、言葉に糖を壊す方向のベクトルが含まれる。従って同意語であるEM経路と記述するが、一部の菌の持つEM系路は糖新生側に向かって駆動することが判明している。例えばIgnicoccus hospitalis、彼らは嫌気的な独立栄養条件下では、ブドウ糖があってもこれを資化しないにもかかわらず、グリコーゲンやデキストリンなどの多糖類を生合成する能力を持っている。Hyperthermus butylicus やPyrolobus fumarii、彼らはブドウ糖を生合成しないだけでなく、多糖類を生合成する能力も持たない。しかしG-6-Pを通ってG-1-Pまで変換する酵素はもっている。
https://www.genome.jp/kegg-bin/show_pathway?select_scale=1.22&query=&map=hbu00010&scale=1.22&orgs=&auto_image=&show_description=hide&multi_query=
これらの細菌のEM系路をどのように捉えるのか。これらの細菌が、カール・ポパーが言うところの蓄積主義の崩壊−白いカラス−になり得るのだろうか。カラスは黒いという無数の蓄積された科学的命題は、1羽の白いカラスの出現で崩壊する。前回述べたように、古細菌の中で主要なグループであるEuryarchaeota属、Crenarchaeota属のいずれにも、グルコースを代謝できない菌は少なくない。ここで言う代謝とは生合成と分解をさす。それは古細菌のことだろうと言われるかもしれないが、そうでもない。真正細菌つまりバクテリアにおいても、Polynucleobacter属やBordetella属をはじめとしてかなり広範囲の菌が、グルコースの生合成系のみならずグルコ−スのリン酸化酵素を持っていない。まあBordetella属細菌は百日咳の病原体で寄生性細菌であるためグルコースまでの代謝を失っている可能性は否定できないが、Polynucleobacter属の Polynucleobacter duraquaeはオーストリアのアルカリ性の湖から単離された自由生活性細菌である。これらの例外が多数知られているにもかかわらず、「EM系はグルコースを分解する経路である」あるいは「逆行するEM系を通ってグルコースが生合成される」とする言明が成立するとは思えない。白いカラスが数知れないほど乱舞しているのである。白いカラスはカラスではないとして、漫然と教科書に従った講義を続けていいのだろうか、と長い間迷っていた。
結論を言えば、我々よりはるかに長い時間を生き続けてきた微生物には、グルコースなど関係なく生きることのできる多くの種が存在するのである。そうであるなら、彼らの持つ代謝系を基盤にした認識体系が併存してもおかしくはあるまい。(注:近年、アルコール中毒患者が更生施設に収容された後で急激に痴呆が進む原因が、これらアルコール中毒患者の脳においてエネルギー源がブドウ糖ではなく酢酸となっているという報告がある。ひょっとすると、これらの患者の脳においては遠い過去に働いていた代謝系が再現されているのかも知れない)
勿論、世の識者がこの程度のことをご存じであることは、承知の上での議論である。教科書なんて、物事が明らかになってその評価が確定する頃にならないと改訂されない代物である。そんな時代遅れのものを相手に議論を吹きかけるのが愚かな行為であることは、百も二百も承知の上で話をしている。多くの研究者と呼ばれる人々は、目の前の競争が忙しくてこんなバカバカしい議論をしている暇はないのであろう。パラダイムが変われば、新たなパラダイムに乗り換えて研究をすればよい。それがコストパフォーマンスの高い効率的な研究法である。論文になりそうにない、自らが依拠しているパラダイムの足下を掘るような仕事は、研究者人生にとってかなり危険な試みになるに違いない。功利的に考えればその通りである。しかしながら、それでいいのかと考え続けてきた。表面的な研究活動は行ってきたが、思考時間の殆どの部分は、役に立たないことに費やされていた。自覚してやってきたのだから、後悔はない。
仕事を辞め、実験化学者としてはすでに終了した私であれば、危険な勇み足になることは承知の上で論を進めても良いだろう。いわゆるEM系路と糖新生系からグルコースを除いた時、どのような風景が見えるのか。
生物の発生時の特徴を多く残していそうな生物群は、好熱性で嫌気的な生物であると考えられている。そのような特徴を持つ生物は、火山の噴気口、特に海底の熱水噴出口周辺に、いまも棲息している。こうした我々から見ると過酷に思える環境から得られた細菌類(バクテリア、古細菌に係わらず)にあって、グルコ−スやグリコーゲンまで達していないがエネルギー消費系であるEM経路(糖新生系)は機能しているようだ。中村 運氏は3-ホスホグリセリン酸からピルビン酸に連なるEM経路の一部を原始発酵系と呼び、原初の生物においてはこの部分の代謝が必要なエネルギーを供給していたように述べているが、私はこの部分の反応のベクトルは逆でありエネルギー消費系であったと考える。では、その反応のエネルギーと担保していたのは何か。
ほとんどの生物でEM経路と同等以上に充実している代謝系がある。ピルビン酸を中心として動いているPyruvate metabolismと呼ばれている系である。この系はピルビン酸とアセチルCoA中心に機能している経路で、この辺りの経路がATPの供給源であろう。今更、生物発生時のエネルギー源論争に乱入する気はないのだが、地中から吹き出すポリリン酸をエネルギー源としていたと考えるのが妥当と考える。アーサー・コーンバーグ博士が見つけた、ポリリン酸を基質としてAMPやADPからATPの生産を触媒するポリリン酸キナーゼのグループは、そういう意味でとても面白い。もっと興味深いのは、広島大学の黒田博士が研究室紹介に書いているATPの起源に関する一文である。一寸紹介するが、ポリリン酸をMg イオンの存在下に加熱すると、3分子の正リン酸が環状の無水物となったトリメタリン酸が優先的に生成するそうだ。このトリメタリン酸がアデノシンと反応すれば、1段階の反応でAMPやADPを経由することなくATPが作られることとなる。これが、ATPが生物の普遍的エネルギー通貨として使われるようになった理由かもしれないと言う話だ。とにかく、6単糖へ続くEM経路の位置づけは、エネルギー産生系ではなくエネルギー消費系に変わってしまう。
ではなぜ、エネルギーを消費してまでG-6-Pまでの生合成系が必要かという問いに答える必要があるだろう。G-6-Pから出発するペントースリン酸経路は、細胞内で使用される還元剤であるNADPH2+の生産を担うとともに、トランスケトラーゼ、トランスアルドラーゼと呼ばれる酵素群による糖の相互変換を通して、核酸の原料となるリボースの生産を担うとされている。この部分の説明にも意義はあるが、それはペントースリン酸経路の項で説明しよう。この場で一言付け加えておくとすれば、ペントースリン酸経路は解糖系の側路という捉え方は間違いであろうし、この経路を6回まわると,グルコース-6-リン酸は全てCO2とNADPH2+に変換されるとする説明は、綿密に計算されたことは認めるにしてもあまりにも発想が貧困であるように感じる。
少し異なる視座から、解糖系について考えてみよう。いわゆる一般的に語られる解糖系において、グルコースからグルコース-6-リン酸、果糖-6-リン酸から果糖-1、6-ジリン酸への変換において2ATPの消費がおこる。果糖-1、6-ジリン酸が2分子の3炭糖(3-ホスホグリセルアルデヒドと13-ジヒドロキシリン酸)へ解裂された後、ピルビン酸までの代謝に伴って4分子のATP生産が起こる。従って消費された2分子のATPを差し引いて正味2分子のATPが生産されると説明される。身も蓋もない言い方をすれば、2分子のATPを投資すると4分子のATP生産が起こるため、差し引き2分子のATPが収益ですよと言っているわけだ。これではまるで資本主義の解説ではないかと思うのだが、妙に説得力があるのも事実である。この説得力こそが、我々が資本主義の世界に住み、その考え方にどっぷりと浸っている証拠であろう。考え方の善し悪しを云っているのではない、その考え方に無意識に溺れていることを指摘しているだけである。
さらに、取り込んだグルコースが分解されることによって生存に必要なエネルギーが発生するという説明で納得する精神構造は、我々が動物であり食物を摂取・分解することで生きているという実感と親和性が高いようだ。従属栄養生物であるというヒトの存在様式に、我々の意識が拘束されていると考えて良い。社会科学的概念を安易に自然科学に持ち込むことには注意深くあらねばならないとは思うが、この親和性の高さはカール・マンハイム云うところの「意識の存在非拘束性」に対応するのではないだろうか。彼は、思想の存在被拘束性を越えて真理に近づくためには、全体的視野から相関や歴史を見よ、いくつもの視座の間を自由に浮動させることが出来る知識人になれと説いた。常識的解糖系の解釈においては、まさに視座の固着が起こっているように感じている。
何度も書くようだが、グルコースなんてEM経路の外につき出した蛇足に過ぎないなどという講義は、さすがに出来なった。学生がこの意見を盲信した場合、彼にとって就職や進学で不利になる場面が予想できるからである。言いたいことをストレスなく言える場所が欲しかったと今でも思っている。