
解糖系についてもう書きたいことは書き終わっただろうと思われるかもしれないが、30年以上、とぼとぼと一人で考え続けてきたことはこれだけではない。解糖系よりも糖新生系の重要さを主張する仮説を補強するために、いわゆる解糖系の周辺回路について、少し補足しておくことにする。
エントナー-ドウドロフ経路(ED経路)は好気性のバクテリアや古細菌でよく見られ、解糖系の一種として認識されている。図11に示すように、この系においてはグルコースがグルコース-6-リン酸に変換された後、グルコース-6-リン酸デヒドロゲナーゼにより基質レベルでの酸化を受け、6-ホスホグルコノラクトンとなる。6-ホスホグルコノラクトンは加水分解をうけて6-ホスホグルコン酸となった後、ホスホグルコン酸デヒドラターゼにより脱水反応に続くケト型への異性化を起こし、2-デヒドロ-3-デオキシ-6-ホスホグルコン酸を与える。2-デヒドロ-3-デオキシ-6-ホスホグルコン酸はレトロアルドール縮合を起こして、3-ホスホグリセルアルデヒドとピルビン酸ヘと変換される。3-ホスホグリセルアルデヒドは先に述べた解糖系と同じ経路を通って、2分子のATPを生産しながらピルビン酸となった後、先に生成していたピルビン酸と合流してその後の代謝系へと流入する経路である。
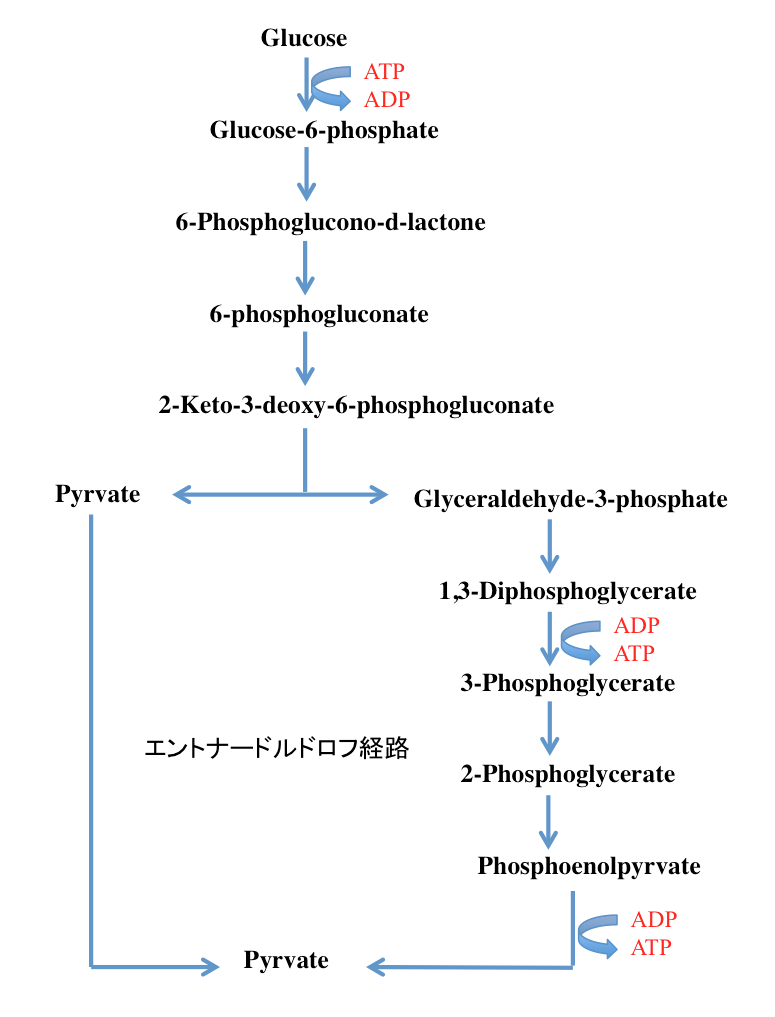
グルコースからのATP収支を考えれば、消費されるATP は1分子、生産されるATPは2分子となるため、グルコースからピルビン酸までの代謝においては1分子のATPが生産されることになる。NADH2+の収支では、通常のグリコリシスと同じく2分子のNADH2+が生合成される。この系に対し、「ATPの収支においてはグルコース1分子当たりATP1分子とEM経路よりも少なく、系が単純な分やや効率は悪い」、あるいは「基質レベルでのリン酸化は片系列でしか行われず ATP の生成は 「EM 経路」 の半分しかない」と、ATP生産量を基準として劣った効率の悪い系として説明される場合が非常に多い。
さらに、いくぶん言い訳っぽく聞こえるが、「発酵としてのエネルギー効率は EM 経路より悪い訳だが、この経路を所持しているのは発酵菌ではなく好気性細菌で、この経路を使う主目的は糖を酸化しやすいピルビン酸に転換するところにあると思われる」という内容の文章を付け加えて、TCA回路に続く酸化的リン酸化を考慮すれば、それほどの損失ではないと説明される場合も見受けられる。そのような解釈で本当に良いのだろうか?
もう一つ、解糖系の仲間に触れておこう。超好熱性古細菌Pyrococcus furiosusはピロ解糖系と呼ばれる少し変わった解糖系を持つ。この系においては、6単糖段階でのリン酸化を経由せずに3炭糖であるグリセルアルデヒドとピルビン酸への解裂を起こす。
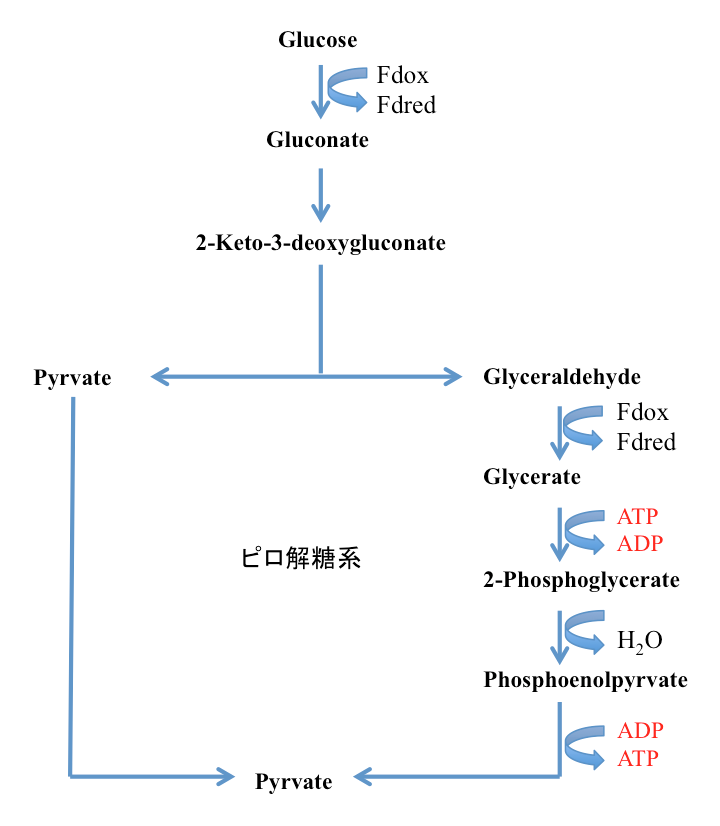
図12に示すように、グルコースが脱水素を受けてグルクロノラクトンとなった後、脱水、エノール型からケト型への異性化を起こして、2-デヒドロ-3-デオキシ-グルコン酸が生成する。2-デヒドロ-3-デオキシ-グルコン酸の3位と4位の炭素間結合が、レトロアルドール縮合を起こして解裂し、ピルビン酸とグリセルアルデヒドを生成する系である。グリセルアルデヒドはグリセリン酸へと酸化された後、グリセリン酸-2-キナーゼによって2-ホスホグリセリン酸となり解糖系に合流する。ホスホエノールピルビン酸からピルビン酸への段階において1分子のATPの生産が起こるが、グリセリン酸から3-ホスホグリセリン酸への段階で1分子のATPが使われているため正味のATP生産は起こらない。
これら二つのの系路と分布する生物を比較して、高等な生物はATPの生産効率が高くなるように進化したと誘導する説明をみかけることがあるが、この説明は生物の生息環境や系の駆動方向を軽視したところで成立する幾分以上にアドホックな説明であろう。そして、そのアドホックな判断基準の根源には、現在の我々を覆っている効率第一主義があると考えている。企業において、コストパフォーマンス、生産効率、エネルギー効率を求めることは理解できないこともないが、教育分野にこれを持ち込まれると、私は頭を抱えてしまう。教育現場でPDCAサイクルを高速で回せといわれ始めたら、教育は崩壊するに違いない。とにかく,現代人は効率化という概念に縛られすぎているように思う。そして、その価値基準をもってATPの生産効率が高い生物の方がより進んだ生物であると決めつけているように思われる。あまり望ましくない「意識の存在被拘束性」の一例だろう。
今となってはちょっと古くなってしまったが、図13は1992年Schaefer, T. and Schoenheit, P.の報告において使われている超好熱性古細菌 Pyrococcus furiosus のピロ解糖系である。図の最下段部分で、Acetyl-CoAからAcetateになる部分でATPとADPを結ぶ矢印の方向が間違っているのでそのつもりで見ていただくとして、彼らはピロ解糖系の話をしているにもかかわらず、ピロ解糖系に含まれないピルビン酸から酢酸に連なるATP生産系を図に書き加えている。
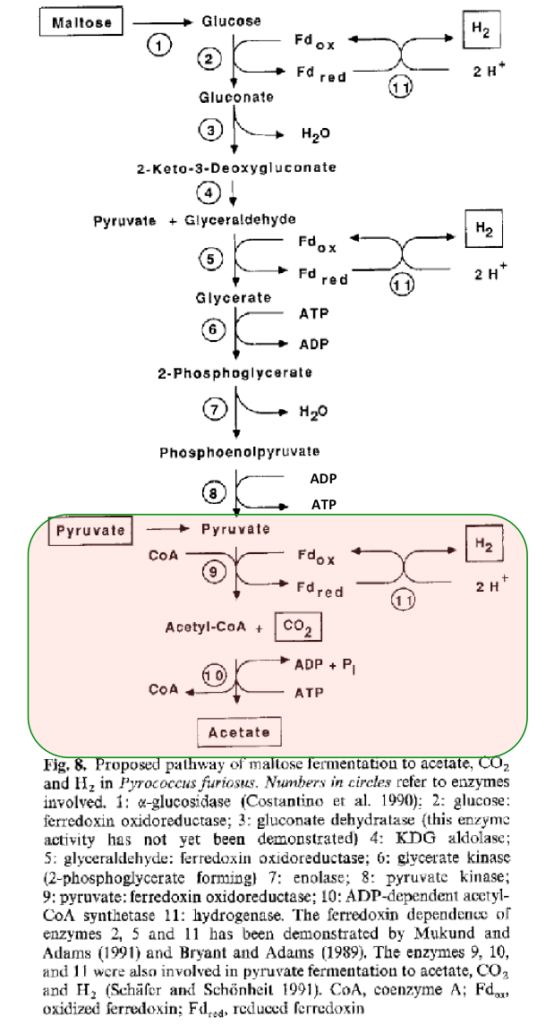
Schaefer, T. and Schoenheit, P. Microbial.(1992) 188-202
先に述べた説明において、解糖系に分類される3種の系路の価値判断基準は、ATPの生産効率にある。つまり、解糖系においては、ATPが生産されなければならないとする固定概念が存在しているだけでなく、この概念の縛りはとても強いようだ。上述したPyrococcus furiosusの持つピロ解糖系について書いてあるいくつかの総説等には、以下のような記述がある。
《1. 経路を通してATPの再生反応が成立しており、ATPを新たに生成しないことである。エネルギー獲得という解糖の生理的意義を考えると、非効率的な経路であると言える: 蛋白質・核酸・酵素 56(6)、 351、 2012》
《2. 特徴的な点は、グルコースがピルビン酸に酸化される際ピリジンヌクレオチドは関与せずフェレドキシン関与であること、しかも発生する還元力は全て分子状水素の生成に使われてしまいATP合成とは共役していないこと、そして、ピルビン酸の発酵により水素、二酸化炭素、酢酸が生じるが、この際基質レベルでのリン酸化によりATPが生産されることである。全体ではグルコース1分子から2分子のATPが生産されることになる。: 蛋白質・核酸・酵素: 38(10)、 1575-1578、 1993》
《3. ・・・しかしながらP. furiosus はH2ガス圧の高い条件下で培養すると発酵産物を酢酸からアラニンに変更することが知られており、この場合、ピルビン酸からアラニンが生成する間にはATP産生部位が存在しないので、従来のモデルでは正味のATP生産がゼロになってしまうという矛盾が生じる。・・・このことは、本菌においてグルコースからピルビン酸が生成する間のATP収支がゼロではないことを示唆している。蛋白質・核酸・酵素 48(9)、 1256-1262、 2003》
私がこれらの考察に対して抱く違和感は、Pyrococcus furiosusが海底にあるイオウ熱水噴出口から分離された偏性嫌気性超好熱古細菌であり、グルコースが摂取できないような所に生息していることを無視していることに起因する。確かにPyrococcus furiosusは、従属栄養的に培養が可能である。だがその際、マルトース、セロビオース、デンプン、β-グルカンは速やかに資化されるが、グルコース、フルクトース、ラクトース、ガラクトースなどの単糖類はほとんど利用されないことが知られている。にもかかわらず、上記の説明ではグルコースを出発点としたATP収支を議論している。さらに解糖系の終点はピルビン酸という定義があるにもかかわらず、酢酸までこれを延伸してATP生産を議論するのは、恣意的な系の改変だと思う。何故グルコースを系の出発点に置くのか、何故ATP生産に拘るのか、何故酢酸までの系が付け加えられるのか、理解しがたい。自然状態におけるこの菌の変形EM系路のベクトルは、通常云われているEM系路と逆のベクトルを持つ糖新生であり、かつATP消費系であると思うのだが?
解糖と糖新生という代謝系を歴史的視座から見た場合、解糖系を、「真核生物であれ原核生物であれ、ほとんどの生物が持つ、嫌気的でもっとも普遍的でもっとも根源的な代謝系」として認めることはできない。「糖新生系こそが、真核生物であれ原核生物であれ、ほとんどの生物が持つ、嫌気的でもっとも普遍的でもっとも根源的な代謝系」であり、その後に現れた従属栄養生物において、先行する生物が生合成した糖類を、糖新生とは逆向きに動かしてエネルギーを獲得する解糖系が成立した。」とするのが正しい理解であろう。
こうした考察に通底しているのは、EM経路はATP生産系路でありより多くのATPを生産する経路が優れているというパラダイムである。上記2が引用している1992年Schaefer, T. and Schoenheit, P.の報告において、彼らが、解糖系ではATP生産が起こるべきだとするパラダイムの圧力に屈したのかどうかは定かではないが、いわゆるアセチルCoA経路をそっと付け加え、2ATPが生産されるとする図が掲載されている。
さて、この解糖とよばれる系において、出発物質をグルコース、終点をピルビン酸としたのは何故であろうか? Gustav Embden やOtto Meyerhofをはじめとする多くの人々の研究によって1930年代中頃までに解糖系の原型が解明されていたわけだが、系の出発物質に関しては彼らも迷ったのではないだろうか。その迷いが、一寸古い代謝マップにおいて、出発物質をグルコースに限定せずに、グリコ−ゲンからグルコースへの変換系及び グリコーゲンからG-1-Pを通ってG-6-Pへと導く系残していた様に思われる。《以下私見だが、メタン細菌などが糖新生に向かって働くEM経路を持ちグリコ−ゲンを生合成していることを考え合わせると、そのような独立栄養生物においてグルコースを解糖系の出発物質と認めることはなかなか難しい》
今ひとつの疑問だが、Gustav Embden やOtto Meyerhof達のグループは、何故解糖系の終点をピルビン酸としたのであろうか。当時、解糖が起こった際に乳酸が生成することはよく知られていたし、一部の乳酸が グリコーゲンへ再合成されることも知られていた。同じころEduard Buchner、Otto Warburg や Hans von Euler-Chelpin達のグループは、アルコール発酵についての研究を続けていた。エタノールや乳酸を系の終点とすれば、細胞内の還元状態の問題もクリアーに説明できる。しかし、エタノールや乳酸を終点としたのでは、系はアルコール発酵あるいは乳酸発酵になってしまう。それは、筋肉生理をやっていたGustav Embden やOtto Meyerhof達のグループには受け入れがたい結論であったようだ。彼らは、ピルビン酸から連なる別の代謝系の存在を仮定して、恣意的にピルビン酸を終点としたと思われる。
その仮説を満たす化合物である「Acetyl CoA」の発見は、1945年 Fritz A. Lipmannによって達成される。ここで目出度く解糖系とTCA回路の連結が完成したのである。何となくだが、微生物の発酵現象を追いかけていたEduard Buchner、Otto Warburg や Hans von Euler-Chelpin達のグループと、筋肉生理を追いかけていたGustav Embden やOtto Meyerhof達のグループ間に激しい争いがあったような形跡がある。この後にも、何度も見られる農学と医学の軋轢の一例であるのかもしれない
ただし、争いはあったが双方の意見に根源的な矛盾は存在しなかった。これは生化学という学問の黎明期において一つの代謝系を明らかにするという観点から見ると、幸運であったろう。なぜなら、発酵を追っていたにしろ筋肉での代謝を追っていたにしろ、まずグルコースあるいはグリコーゲン(デンプン)があらかじめ存在するという条件下での競争であり、これらを分解してエネルギーを得るという解糖系のベクトルには違いがなかったのである。
しかしながら、解糖という系を考えるに当たって、出発物質をグルコース、終点をピルビン酸、系の意義を嫌気的条件下におけるATP生産系としたことが、先に述べたように今に続く迷走をもたらしたとも考えられる。いや、多くの研究者は迷走しているなんて夢にも思っていない。そんなことを考えているのはごくごく少数だろう。
それはそうとして、とにかく解糖系はグルコースからピルビン酸までと定義された。めでたくAcetyl CoAも見つかって、TCA回路との連結も完成した。目出度し目出度しと言いたい所だが、困ったことにピルビン酸からAcetyl CoAへの変換反応はどちらの系にも属さないことになってしまった。さらにTCA回路は回路であるから、回路の外側にあるAcetyl CoAを含まないとする解釈もある。これでは両系を通してATP収支を考える場合はなはだ不便である。
歴史的経緯を無視できるならば、解糖系の最終物質をAcetyl CoAとするか、ピルビン酸をTCA回路の出発物質として定義した方が、二つの系路の接続が理解しやすい。とはいえ、解糖系はサイトゾル内に存在するため、解糖系をAcetyl CoAまで延伸するのは難しいだろう。同じくピルビン酸は解糖系に含まれているので、これをTCA回路の出発物質とするのも難しそうに思える。
だが、定義などというものは分かり易くかつ使いやすいように決めるのが望ましい。解糖系の終わりはサイトゾル内にあるピルビン酸、TCA回路の開始はミトコンドリア内にある(運ばれた)ピルビン酸としておけば、初学者にとって非常にわかりやすくなると思うのだが・・・。
図14に従属栄養生物、独立栄養生物(光合成をする生物)、独立栄養生物(化学合成細菌)の持つEM経路の基本的な代謝ベクトルを描いてみた。歴史生物学を標榜する人間として発言するとすれば、橙色で示した化学合成独立細菌のもつ経路の出現が最も早く成立したのは間違いない。多糖類や脂質やタンパク質などの生合成を行う微生物が現れた後で、それらの生産物を資源として利用する微生物が出現したと考える。これらの微生物は、新たに生産物の利用系路を作ったのではなく、先に存在した生合成経路を逆行する形で利用系を作ったのであろう。
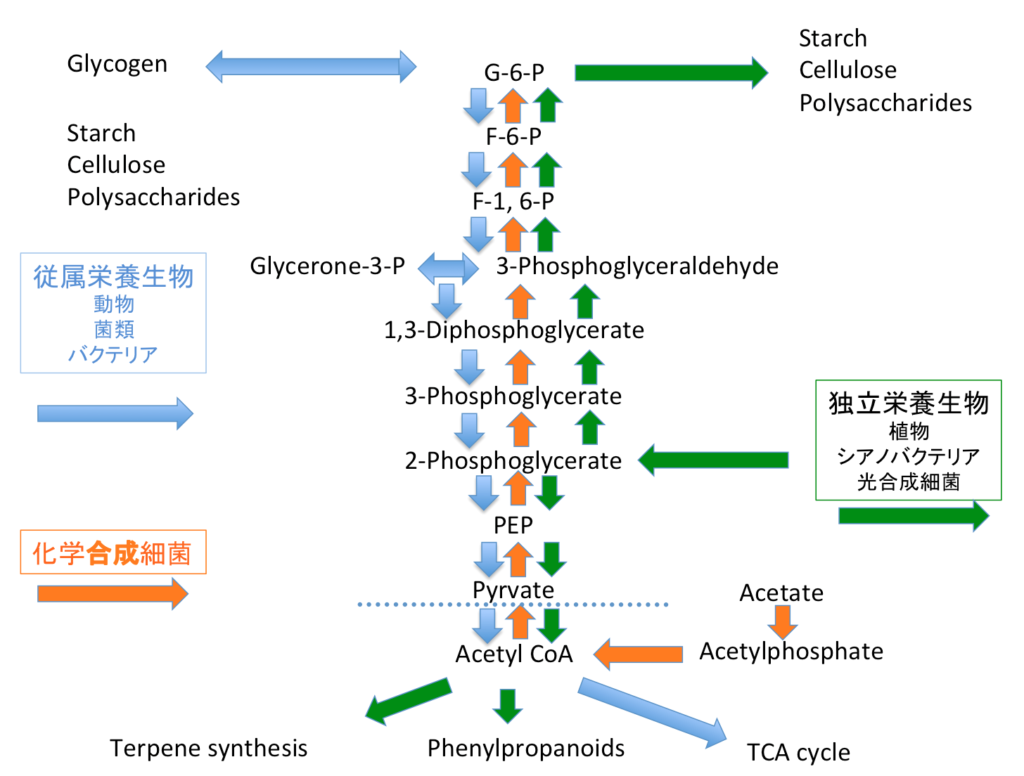
一方、植物やシアノバクテリアなど光合成能を獲得したグループは、すでに成立していたペントースリン酸回路を逆向きに利用して二酸化炭素の固定を行い、無尽蔵とも思えるような量の3-ホスホグリセリン酸の生合成を達成した。彼らもまた、いわゆるEM系路を利用するのだが、3-ホスホグリセリン酸が系への流入物質となるために、彼らの持つEM系路はここから上下に分かれる形での物質変換系として捉えるのが理性的ではないだろうか。
上記のような話をすると、話の途中で「かなり偏向しているね」とか、「独断の傾向が強いな」などと云う賞賛(?)を受ける場合が多い。「そんな考え方もありうるかもね」と、現行のパラダイム側にたって婉曲に否定される場合も少なくない。再度力説するが、植物の生き方に立脚すれば、現行の解糖系についての解釈など妄想に過ぎなくなる。植物生理学の教科書に、現在認められている解糖系の説明を記載すべきではないと思う。
植物は光合成を行う。教科書には、光合成とは太陽光のエネルギーを利用して二酸化炭素から糖とATPと酸素を作る反応であると書かれている。そして、グルコースやショ糖やデンプンがつくられると無造作に書いてある。しかし、そういう理解では、植物に対する洞察がおかしくなってしまう。植物はカルビンベンソンサイクルを動かし、光合成生産物を3-ホスホグリセリン酸の形でEM経路に流入させているのである。この3-ホスホグリセリン酸を起点として、EM経路を逆行した同化産物はグルコース-6-リン酸を通って、デンプン、セルロース、マンナン、アガロースなどの多糖類を形成する。(グルコースへ行くのはほんのちょっぴりである。)同時に、逆向きに回る還元的ペントースリン酸経路の構成成分であるリボース-5-リン酸が、核酸に繋がる代謝系へ流入することになる。
他方、起点である3-ホスホグリセリン酸から順行した同化産物は、アセチルCoAを経由して膨大な量のテルペノイドやフェニルプロパノイドに連なる代謝系へと流入していく。ここにおいて、植物のEM経路の出発物質は3-ホスホグリセリン酸と決めたほうが理性的な判断であると考える。さらに、一寸ばかり意地悪なデータだが、植物にはG-1-PやG-6-Pからダイレクトにグルコースを生産する酵素は持っていない。一寸甘めの植物、砂糖ダイコンもブドウでさえも持っていない。彼らはG-1-PからUDP-グルコースを経由してショ糖を作った後、これを加水分解して果糖とグルコースとしているわけで、ここで作られるブドウ糖はEM経路を逆行した生産物ではない。もちろん、ショ糖を経由するブドウ糖合成系は先に述べた好熱性古細菌などのグル−プには存在しない。解糖系を、「真核生物であれ原核生物であれ、ほとんどの生物が持つ、嫌気的でもっとも普遍的でもっとも根源的な代謝系」として説明に対しては、白いカラスが群舞しているではないか。既存の枠組みにあまりにも固執しすぎているのではないだろうか。
結論だが、解糖と糖新生という代謝系を歴史的視座から見た場合、解糖系を、「真核生物であれ原核生物であれ、ほとんどの生物が持つ、嫌気的でもっとも普遍的でもっとも根源的な代謝系」として認めることはできない。「糖新生系こそが、真核生物であれ原核生物であれ、ほとんどの生物が持つ、嫌気的でもっとも普遍的でもっとも根源的な代謝系」であり、その後に現れた従属栄養生物において、先行する生物が生合成した糖類を、糖新生とは逆向きに動かしてエネルギーを獲得する常識的な解糖系が成立した。」とするのが正しい理解であろう。
まあ読んで理解したとしても、グリコリシスへの見方が少し変わる程度の問題である。そう位置づけて読み捨てても良いかもしれない。私も、上記の内容を構築したからといって、日常の研究活動に影響が出たことはないと思う。しかしながら、生物、特に植物の代謝に対して考察を進めていくに際して、非常に大きな影響を与えることがわかってきた。さらに、次に書くTCA回路に関する現在の解釈の誤りを考え合わせて考えると、生物の持つ基礎的な代謝系の解釈が大きく変わってしまうと考えている。タマネギの収穫は今日終わらせた。ジャンボニンニクの収穫と田植えと農地の草刈り、それに防除作業が一段落したら、解糖系の話を書き始めることにする。要するに解糖系は逆さまに動いていた、そしてTCA回路は回っていなかったという話になる予定である。
近頃、夜更かしの度が過ぎている。寝たのは、一昨日が3時半過ぎ、昨日が2時40分頃、今日ももうすぐ1時である。近所の防犯には役立っているようだ。午前3時半といえば、朝刊の配達の時間である。根室付近だったら夜が明け始めているのではないかな。でも、昼間疲れると夜が眠れないという変な現象が起こってしまう。疲れが累積しそうで気にしているのだが、眠たくないのだからどうしようもない。