
沈思黙考の5、データが古い以前の分をアップしてしまいました。新しいKEGGのデータを基に書き直したのですが、間違って消してしまいました。明日にでも新規なものに入れ替えますので、以下の分は読まないで下さい。
《 だが、生き物というものは一筋縄で理解できるものではない。進化的に見ると、単子葉植物は双子葉植物から分岐して進化してきたようである。進化していることが良いとか悪いとか言う判断はすべきではない。単に単子葉植物の方が進化が進んでいるというにすぎない。そこでだが、ゲノム解析の終わった植物群の中にも、単子葉植物が存在する。イネ(japonica)、ソルガム(モロコシ:年のヒトはコーリャンといった方が分かり易いかも)、トウモロコシ、セイヨウヤマカモジとアワの解析が終わっている。これらの植物についてアブシジン酸生合成系を眺めてみると、困った事実が判明する。下にジャポニカ種のイネの代謝系を占めそう。
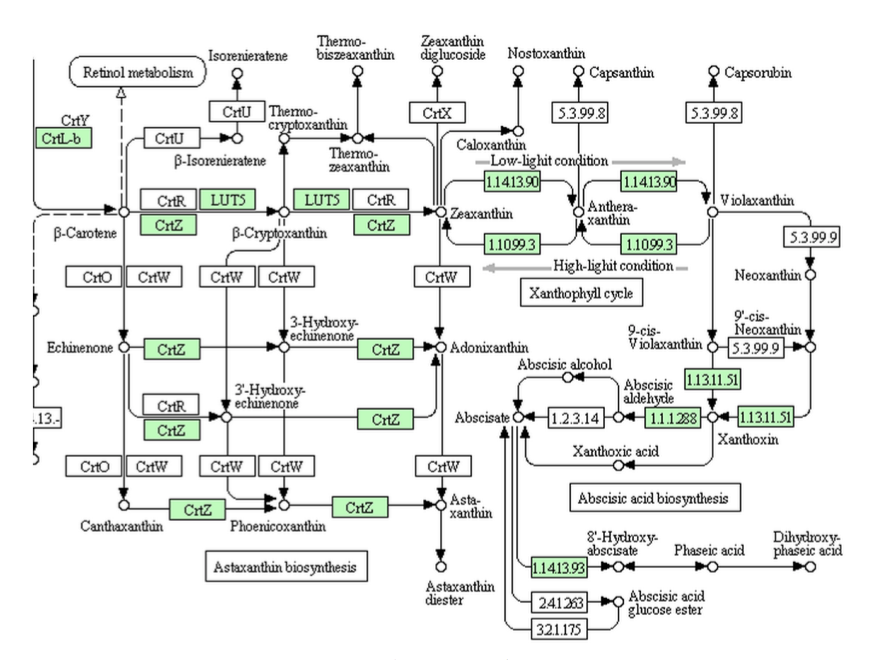
図13-7にイネのデータを示しているが、XanthoxinからABA-アルデヒドまでの代謝系はあるのだが、ABA-アルデヒドをアブシジン酸に導くaldehyde oxidaseが存在しない。アブシジン酸生合成の最終段階にあるはずのaldehyde oxidaseが存在しない?この酵素がないのにアブシジン酸生合成系と言えるのだろうか。
ところが、この段階の酵素を欠いた植物はめずらしくないのである。マメ科のダイズ、タルウマゴヤシ、バラ科のエゾノヘビイチゴ、ウリ科のキューリ、ヤナギ科のポプラ(black cottonwood)そしてナス科のトマトもこの酵素を欠いている。ゲノム解析の終わった蘚類以上の植物18種の中で、この酵素を持つのはシロイヌナズナ、ミヤマハタザオ、トウゴマ、そしてヨーロッパブドウの4種に過ぎない。いろいろな植物でABA生合成系が何処まで機能しているのかをまとめると、以下のようになる。
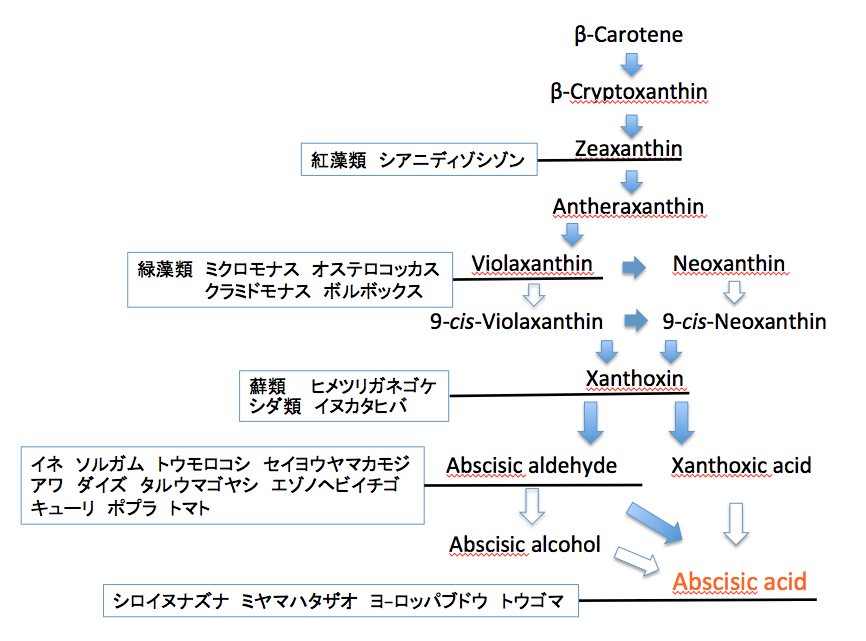
結論だが、β-カロテンを通ってアブシジン酸に達する生合成経路と、これを消去して8-ヒドロキシアブシジン酸にする酵素を併せ持つ植物は、カラシナ科、トウダイグサ科そしてブドウ科にしかみいだせない。この事実を基に、アブシジン酸はストレス存在下に急速に生合成されるとか、ストレスが解除されると速やかに代謝されるなどという一般化した言明は、本当に可能なのであろうか。我々が見ていたのは、色々な植物の持つ代謝系を重ね書きして、一見完成したように見える「苟且の代謝マップ」ではなかったか。》
以下の分は読まないで下さいと書いてはみたものの、読まないはずはないでしょうね。こんな場合、人は読むなといわれれば読むし、見るなといわれれば見る生き物です。鶴の恩返しであっても伊邪那岐命の神話であっても、見るなといわれれば見るものです。従って、上の部分がどう変わったかをこの下に書き加えることにした。書き換えていた原稿の保存を忘れ、編集ページを閉じたのが原因であるため、書き換えた原稿は消えてしまっているので、再度新たに書くことになってしまった。
だが、生き物というものは一筋縄で理解できるものではない。進化的に見ると、単子葉植物は双子葉植物から分岐して進化してきたようである。進化していることが良いとか悪いとか言う判断はすべきではない。単に単子葉植物の方が進化が進んでいるというにすぎない。そこでだが、ゲノム解析の終わった植物群の中にも、単子葉植物が存在する。イネ(japonica)、ソルガム(モロコシ:年のヒトはコーリャンといった方が分かり易いかも)、トウモロコシ、セイヨウヤマカモジとアワの解析が終わっている。これらの植物についてアブシジン酸生合成系を眺めてみると、困った事実が判明する。下にジャポニカ種のイネの代謝系(2013年)を示そう。
ジャポニカ種のイネのデータを示しているが、β-カロテンからXanthoxinを通ってABA-アルデヒドまでの代謝系はあるのだが、ABA-アルデヒドをアブシジン酸に導くaldehyde oxidaseが存在しない。つまり、アブシジン酸生合成の最終段階にあるはずのaldehyde oxidaseが存在しないのである?イネだけでなくソルガム、トウモロコシ、セイヨウヤマカモジ、アワのすべてがこの酵素を欠いている。この酵素がないのにアブシジン酸生合成系と言えるのだろうか。ところがKEGGも生き物であり、常に新しいデータを基に改訂が続けられている。そこでどうなったか。以下に示す。
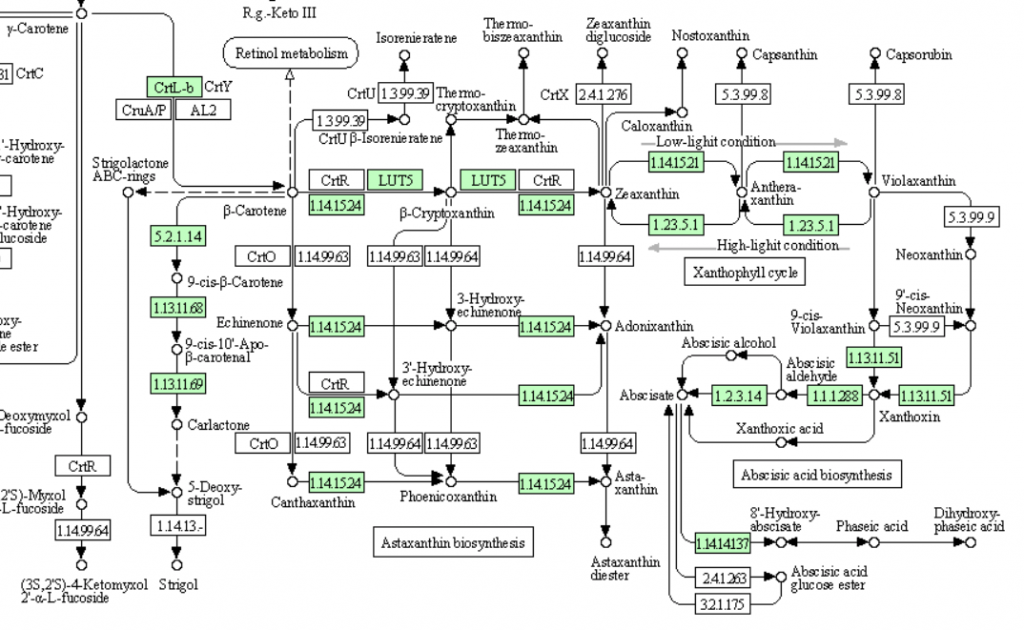
なんと、1.2.3.14(Abscisic aldehyde oxidase)が存在している。つまり、先の議論は成立せず、イネを含むすべての植物は完全なアブシジン酸生合成を持つということになった。まあ、進化学的にもっとも進んでいるという単子葉植物であるので、そうかと思うだけである。だが、2013年時の代謝マップでこの段階の酵素を欠いた植物はめずらしくないのである。(マメ科のダイズ、タルウマゴヤシ、バラ科のエゾノヘビイチゴ、ウリ科のキューリ、ヤナギ科のポプラ(black cottonwood)そしてナス科のトマトもこの酵素を欠いている。ゲノム解析の終わった蘚類以上の植物18種の中で、この酵素を持つのはシロイヌナズナ、ミヤマハタザオ、トウゴマ、そしてヨーロッパブドウの4種に過ぎない。)と書いたのだが、2021年段階においては、これらすべての種に完全なアブシジン酸生合成系が認められていた。
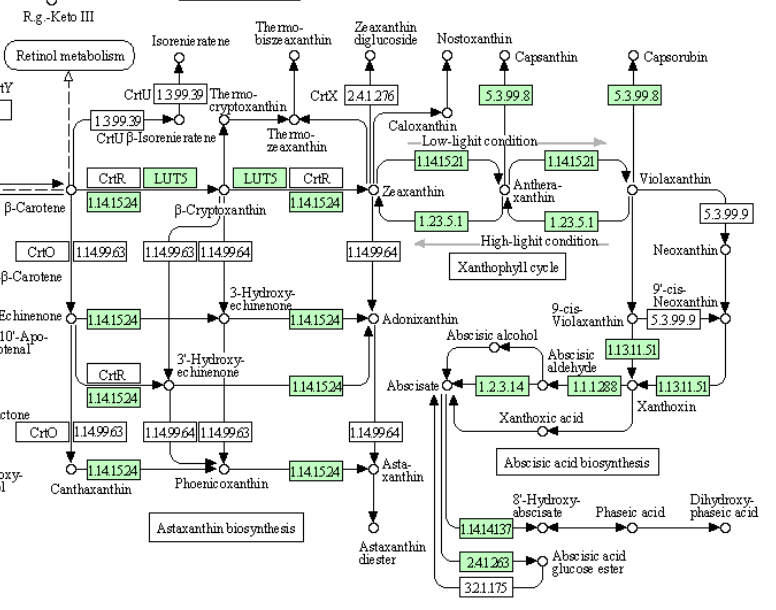
話としてはこの方が合理的であり納得しやすいのだが、少しだけ疑問が残らないでもない。最終段階で働くAbscisic aldehyde oxidaseなのだが、イネにおいては3種の酵素が存在すると書いてある。植物において複数のイソ酵素があること自体は不思議ではない。だが、これらの酵素についてコメント欄を見ると次のように書いている。 While abscisic aldehyde is the best substrate, the enzyme also acts with indole-3-aldehyde, 1-naphthaldehyde and benzaldehyde as substrates, but more slowly. Abscisic aldehydeが最適の基質であるのだが、これらの酵素はインドール酢酸生合成におけるindole-3-aldehyde に対してもゆっくりではあるが働く。生育を抑制するホルモンとされるアブシジン酸の生合成、その最終段階を担う酵素が、成長を促進するインド−ル酢酸(オーキシン)の生合成、その最終段階で働く。いやいやどっちやねんといいたくなる。
少し調べてみると、オーキシン生合成酵素としては別の酵素が比定してあり、その酵素がAbscisic aldehydeに作用するとは書いてないので、Abscisic aldehyde oxidase のインドール酢酸生合成への寄与は少ないのかもしれない。しかしながら、Abscisic aldehyde oxidaseを欠いたままにしておくことへの不安感があったとも考えられる。誤解して欲しくないのだが、これはKEGGがデータを変えたなどという批判ではない。自然界とはそういうものである。もっといえば、こうした議論ができるテーブルを用意していただいたことに、感謝している。
残っている疑問はシダ植物であるイヌカタヒバと蘚類に属するヒメツリガネゴケがアブシジン酸に到達しないABA生合成系しか持たないことかな。それらに対し、アブシジン酸はストレス存在下に急速に生合成されるとか、ストレスが解除されると速やかに代謝されるなどという一般化した言明は成り立つのであろうか。さらに、紅総類と緑藻類においてはアブシジン酸だけでなくルヌラリン酸の生合成も欠いているように見える。緑藻類は陸上植物の直系の祖先とされているとはいえ、水性の藻類だから、水ストレスはないだろうなどという一言で片づけるのは少し難しい。現状では分からないというしかないだろう。結論としていえることは、我々は、色々な植物の持つ代謝系を重ね書きして、一見完成したように見える Reference pathway に基づいて議論をしていただけではなかったか。神は細部に宿る、自戒である。